
Arxius de Miscel·lània Zoològica. Volum 12 (2014) Pàgines: 93-123
Descripción osteológica del rorcual común (Balaenoptera physalus, Linnaeus, 1758) del Museo de Ciencias Naturales de Barcelona
Carrillo, M., Alcántara, E., Taverna, A., Paredes, R., García-Franquesa, E.
DOI: https://doi.org/10.32800/amz.2014.12.0093Descarregar
PDFCita
Carrillo, M., Alcántara, E., Taverna, A., Paredes, R., García-Franquesa, E., 2014. Descripción osteológica del rorcual común (Balaenoptera physalus, Linnaeus, 1758) del Museo de Ciencias Naturales de Barcelona. Arxius de Miscel·lània Zoològica, 12: 93-123, DOI: https://doi.org/10.32800/amz.2014.12.0093-
Data de recepció:
- 10/07/2013
-
Data d'acceptació:
- 16/07/2014
-
Data de publicació:
- 03/09/2014
-
Compartir
-


-
Visites
- 5796
-
Descàrregues
- 592
Resumen
Descripción osteológica del rorcual común (Balaenoptera physalus, Linnaeus, 1758) del Museo de Ciencias Naturales de Barcelona
Se presenta la descripción osteológica del esqueleto del rorcual común (Balaenoptera physalus, L., código de registro MZB 83-3084) del Museo de Ciencias Naturales de Barcelona (MCNB). El espécimen procede de un varamiento en Llançà (Girona) en 1862. El esqueleto pesa 1.161,59 kg y mide 14,6 m, aunque este valor no se corresponde con la longitud total del animal vivo ya que no incluye la longitud del tejido intervertebral. El esqueleto reconstruido, incluido el espesor del tejido intervertebral, alcanza una longitud de 17,35 m. El cráneo pesa 484 kg y tiene una longitud cóndilo-basal de 431 cm que representa el 24,84% de la longitud total y se corresponde con los valores medios obtenidos para otros ejemplares en el Mediterráneo. La columna vertebral posee 58 vértebras estructuradas según la fórmula C7 T14 L14 Ca23 con una longitud total de 10,29 m y un peso de 470,95 kg. Todas las vértebras cervicales están libres y muestran compresión dorsoventral a diferencia de las vértebras torácicas, las lumbares y primeras caudales que son relativamente uniformes, de sección circular. A partir de la Ca14 se produce un cambio en la forma de las vértebras y la relación entre la anchura y la altura es superior a 1, lo que indica una compresión lateral. Ni en las epífisis de la ulna y el radio ni en las de ninguna vértebra se aprecian líneas de sutura, lo que indica que se trata de un ejemplar adulto que ha alcanzado la madurez en su proceso de osificación.
Palabras clave: Rorcual común, Esqueleto, Osteometría, Mediterráneo
Abstract
Description of the skeleton of the fin whale (Balaenoptera physalus, Linnaeus, 1758) at the Natural History Museum of Barcelona
We describe the osteology of the fin whale (Balaenoptera physalus, L., registration code MZB 83-3084) at the Natural Science Museum of Barcelona (MCNB). The specimen was stranded and died on a beach in Llançà (Girona, Spain) in 1862. The skeleton weighed 1, 161.59 kg and measured 14. 6 m, although the live animal would have been longer because the invertebral tissue was not included in the length originally stated. The newly reconstructed skeleton includes the invertebral discs and is 17.5 m long. The skull weighs 484 kg and the condilobasal length is 431 cm ( 24. 84% of the total length), corresponding to the mean length of specimens in the Mediterranean. The vertebral column has 58 vertebra structured following the formula: C7 T14 L14 Cd23. It is 10.29 m long and weighs 470.95 kg. All the cervical vertebra are free and show dorsoventral compression, differing from the thoracic, lumbar and first caudal vertebra that are relatively uniform, and rounded. From Cd14 se onwards, the vertebra vary in shape and the relationship between width and height is greater than 1, indicating lateral compression. The lack of suture lines in the epiphysis of the ulna and radius indicates the specimen is an adult that has reached ossification maturation.
Key words: Fin whale, Skeleton, Osteometry, Mediterranean
Resum
Descripció osteològica del rorqual comú (Balaenoptera physalus, Linnaeus, 1758) del Museu de Ciències Naturals de Barcelona
Es presenta la descripció osteològica de l’esquelet del rorqual comú (Balaenoptera physalus, L., codi de registre MZB 83-3084) del Museu de Ciències Naturals de Barcelona (MCNB). L’espècimen procedeix d’un encallament a Llançà (Alt Empordà) l’any 1862. L’esquelet pesa 1.161,59 kg i mesura 14,6 m, tot i que aquest valor no es correspon amb la longitud total de l’animal viu atès que no inclou la longitud del teixit intervertebral. L’esquelet reconstruït, incloent-hi el gruix del teixit intervertebral, assoleix una longitud de 17,35 m. El crani pesa 484 kg i té una longitud condilobasal de 431 cm que representa el 24,84% de la longitud total i que es correspon amb els valors mitjans obtinguts per a altres exemplars al Mediterrani. La columna vertebral consta de 58 vèrtebres estructurades segons la fórmula C7 T14 L14 Ca23 amb una longitud total de 10,29 m i un pes de 470,95 kg. Totes les vèrtebres cervicals estan lliures i mostren compressió dorsoventral a diferència de les vèrtebres toràciques, les lumbars i les primeres caudals que són relativament uniformes, de secció circular. A partir de la Ca14 es produeix un canvi en la forma de les vèrtebres i la relació entre l’amplada i l’altura és superior a 1, indicativa de compressió lateral. Ni a les epífisis de l’ulna i el radi ni a les de cap vèrtebra s’aprecien línies de sutura, la qual cosa indica que es tracta d’un exemplar adult que ha assolit la maduresa en el procés d’ossificació.
Paraules clau: Rorqual comú, Esquelet, Osteometria, Mediterrani
Introducción
La primera referencia publicada sobre este rorcual proviene de la Llibreta de Memòries de Joan Baptista Serinyana (1818-1903), un campesino vecino de Llançà (Girona) que relató el varamiento del rorcual vivo en la punta de Cap Ras el 16 de junio de 1862. Cuatro días más tarde, el 20 de junio, se publican en la prensa de Cataluña (El Telégrafo y Diario de Barcelona de Avisos y Noticias) unas notas sobre el varamiento del rorcual, con las primeras referencias sobre sus dimensiones y peso. Según estas noticias, la ballena mediría 17,35 metros y pesaría 23 toneladas, aunque en otra noticia publicada más adelante el peso que se da es de 900 quintales, que equivaldría a 41 toneladas, y tan solo la lengua pesaría 874 kilos. Entre las numerosas personas que acuden a Llançà a ver la ballena varada se encuentran algunos visitantes ilustres, como Narcís Monturiol (1819-1885), inventor de un precursor del submarino, y Paul Gervais (1816-1879), eminente zoólogo. El primero se interesó por conocer el volumen de los pulmones y el segundo compró un ojo y el corazón, interesándose también por adquirir el esqueleto completo. La Universidad de Barcelona se interesó por la ballena y el rector, Víctor Arnau, designó para adquirir el esqueleto al catedrático Dr. Julián Casaña y Leonardo, que se desplazó hasta Llançà acompañado de José Boix, director del Instituto de Figueres. Finalmente, el 1 de julio se cerró el acuerdo de compra con los pescadores que habían recuperado y limpiado el esqueleto de la ballena y se procedió a su embarque rumbo a Barcelona.
El 18 de julio de 1862 el esqueleto se trasladó a la Universidad de Barcelona y se terminó de montar en septiembre de 1881. El montaje lo dirigió Francisco Vidal, ayudante de la cátedra de Historia Natural de la Universidad de Barcelona, bajo la dirección del decano de la Facultad de Ciencias, José Planellas Giralt. El esqueleto montado permaneció expuesto hasta 1917. Ese año se desmontó el museo de la Universidad y el esqueleto de la ballena se trasladó al Museo Martorell. Al esqueleto de rorcual común se le asignó el número de registro 412 (818) en agosto de 1917. En 1925, el esqueleto se trasladó desde el Museo Martorell al Castell dels Tres Dragons, al pasar este a ser la sede del Museo de Zoología de Barcelona. En este edificio permaneció expuesto al público hasta el cierre de las exposiciones en mayo de 2010. El 16 de junio de 2010, fecha en la que se cumplían 148 años de su varamiento, durante el desmontaje se produjo la caída y rotura del neurocráneo. Pero en mayo de 2011 gran parte de las múltiples fracturas del cráneo habían sido ya restauradas en el laboratorio del Instituto Catalán de Paleontología. Las restauradoras habían desengrasado y limpiado el esqueleto completo además de unir y reforzar los fragmentos rotos de los huesos del cráneo. No obstante, quedaban aún por unir cuatro grandes piezas, las maxilas y premaxilas, cuyo montaje finalizó el Gabinete de Estudios Ambientales (GEA). Es conveniente hacer hincapié en el delicado estado de conservación del espécimen, resultado de su reciente historia y también de su antigüedad, lo que, unido a su valor patrimonial, obligó a que su manipulación y montaje se llevaran a cabo con extremo cuidado.
La colaboración establecida entre el MCNB, GEA y Canarias Conservación permitió reunir un equipo multidisciplinar que durante dos meses terminó la reconstrucción del cráneo y realizó el montaje del esqueleto en el Castell dels Tres Dragons (parque de la Ciutadella). A finales de junio de 2011, el esqueleto del rorcual salió reconstruido y parcialmente montado hacia la nueva sede del Museo de Ciencias Naturales, en el Fòrum, acabando una estancia de 86 años en el edificio y de 64 años en la emblemática Sala de la Ballena.
Material y métodos
De forma preliminar a la descripción osteológica del espécimen, todos los huesos fueron examinados para evaluar su estado de conservación señalando aspectos como pérdidas de hueso, fracturas o deformaciones. Se fotografiaron todos los huesos, tanto por su parte anterior como en vista lateral. Las imágenes cuentan con una escala en centímetros y una etiqueta para su identificación. Con esta información se elaboró un archivo osteográfico en el que quedó registrada la totalidad de las piezas óseas que componen el esqueleto del espécimen.
En cuanto a las medidas generales del esqueleto, se siguieron los criterios utilizados por Perrin (1975) y los de Buchholtz & Schur (2004) para las medidas del esqueleto vertebral. Para la descripción del cráneo se utilizaron las pautas de Wada et al. (2003) en su descripción de Balaenoptera omurai.
Cada vértebra fue medida de la siguiente manera: la anchura (CW) y altura (CH) de la superficie del centro se tomaron en la cara anterior de la vértebra y la longitud (CL) se midió en la parte ventral de cada vértebra (fig. 1). El proceso neural (NPH) se midió como la distancia vertical entre la punta dorsal de la espina neural y la línea horizontal del dorso del cuerpo vertebral. La altura del arco neural (NAH) se midió como la distancia entre la metapófisis y la línea horizontal a nivel del cuerpo vertebral (fig. 1). La anchura de la apófisis transversa (TPW) fue medida en el lado izquierdo de las vértebras. Las medidas fueron registradas en centímetros con cinta métrica y calibrador.

Fig. 1. Principales medidas registradas en las vértebras y en el esqueleto craneal (Balaenoptera physalus, MZB 83–3084). Definiciones en tabla 1.
Fig. 1. Main measurements recorded in vertebra and skull (Balaenoptera physalus, MZB 83–3084). Definitions in table 1.
La longitud del esqueleto poscraneal se midió como la suma de la longitud de cada vértebra incluyendo las epífisis. Este valor no es el mismo que el de la longitud poscraneal del animal vivo debido a que no incluye la longitud del tejido fibrocartilaginoso de los discos intervertebrales ni los tejidos de recubrimiento de las porciones distales del esqueleto.
Para la descripción osteológica de la columna se analizó la forma de cada una de las vértebras según la relación entre la longitud (CL), la anchura (CW) y la altura (CH) del cuerpo vertebral. Este parámetro sirve para cuantificar si el centro aumenta proporcionalmente en sus tres dimensiones o, por el contrario, está primando la longitud, por lo que este parámetro se denomina longitud relativa del centro (Buchholtz & Schur, 2004; Buchholtz & Wolkovich, 2005). La longitud relativa del centro de las vértebras implica una variación en el movimiento entre vértebras adyacentes y, por lo tanto, es un indicador de la elasticidad o rigidez de la columna. Así, suponiendo un ángulo de rotación constante, cuando se incrementa la longitud del centro se incrementa el desplazamiento absoluto de la cara posterior de la vértebra. Un CL mayor implica para la columna una menor rigidez y un desplazamiento absoluto mayor (Buchholtz & Schur, 2004). La relación entre la anchura CW y la altura CH nos da la siguiente información: CW > CH indica una compresión dorsoventral. CW = CH superficie de la vértebra con forma circular. CW < CH indica una compresión lateral.
Las costillas que se midieron fueron la primera y la sexta izquierdas, por ser la sexta la mayor costilla de todas. En ambas se registró la máxima longitud y máxima anchura en su parte distal. Las medidas tomadas del esternón fueron también la máxima altura y la máxima anchura. Los huesos de la cintura pectoral fueron medidos en su longitud máxima y anchura máxima en la parte distal a excepción de los huesos de la mano, en los que solo fue medida la anchura transversal de la línea proximal de los carpales. En la escápula se midió la longitud y la anchura de los procesos acromion y coracoides.
Los huesos del esqueleto se pesaron y midieron de forma individual. Para las grandes piezas se utilizó un dinamómetro digital con un polipasto suspendido del techo del taller y, para piezas medianas y pequeñas, balanzas digitales.
Antes de montar el esqueleto se fabricaron 50 discos intervertebrales, para que el montaje definitivo se correspondiera con las medidas reales del esqueleto.
Resultados
Las longitudes parciales de cada parte del esqueleto, así como los pesos de cada una, se encuentran en la figura 2. La longitud total del rorcual sin considerar el tejido intervertebral es de 14,61 m. El conjunto de huesos de todo el ejemplar tiene un peso de 1.037,57 kg. La longitud total conocida a pie de playa (17,35 m) y la del esqueleto limpio (14,61 m) tienen una diferencia de 275 cm, lo que equivale al 15,8% menos de lo medido en fresco. En el montaje para determinar la separación entre las vértebras que correspondería al animal vivo se tuvo en cuenta la longitud estimada a pie de playa, por lo que el espécimen montado alcanza una longitud similar a la que tendría la ballena viva.

Fig. 2. Longitudes y pesos de las diferentes secciones del esqueleto del rorcual Balaenoptera physalus, MZB 83–3084.
Fig. 2. Lengths and weights of the different sections of the whale skeleton, Balaenoptera physalus, MZB 83–3084.
Esqueleto axial
Cráneo
Las medidas tomadas en el cráneo se encuentran en la figura 1 y en la tabla 1. El cráneo del espécimen MZB 83–3084 es relativamente ancho, con una longitud cóndilo-basal de 431 cm que representa el 24,84% de la longitud estimada total del rorcual a pie de playa (17,35 m). La máxima anchura, en la parte postorbital del escuamosal, es de 202 cm. La longitud del rostro es de 301 cm con una anchura máxima de 122 cm y con el margen lateral cóncavo en su mitad posterior, siendo más plano en su mitad anterior. El proceso ascendente de la maxila (PAM) se va ensanchando hacia el extremo posterior. El frontal está expuesto como un estrecho cinturón entre el proceso ascendente de la maxila y el supraoccipital. La inserción de la premaxila con el frontal se encuentra casi en línea con el proceso ascendente de la maxila, llegando a tocar con el frontal. La parte dorsal del proceso supraorbital del frontal está cubierto por el parietal, aunque no es visible en vista dorsal. Las hemimandíbulas tienen una longitud máxima de 418 cm en la izquierda y una altura máxima de 46,5 cm. Cada una de ellas presenta entre seis y nueve forámenes correspondientes a la rama del trigémino, las arterias y las venas de las mandíbulas. El peso total del cráneo es de 484 kg (fig. 2). En la lámina 1 se muestra la fotografía del cráneo completo.

Tabla 1. Medidas en cm del esqueleto del espécimen Balaenoptera physalus, MZB 83-3084. Table 1. Measurements in cm of the skeleton of specimen Balaenoptera physalus, MZB 83-3084.

Lámina 1. Esqueleto craneal del espécimen MZB 83–3084 Balaenoptera physalus.
Plate 1. Skull of specimen MZB 83–3084 Balaenoptera physalus
Aparato hioideo
Los estilohioides derecho e izquierdo son simétricos y tienen una longitud y anchura máxima de 45 cm y 11,5 cm, respectivamente. El basihioides y los tirohioides están fusionados y no se aprecian líneas de sutura. La anchura máxima del hioides es de 86 cm y la altura máxima de 28,5 cm (tabla 1). El peso total del aparato hioideo es de 4,7 kg (fig. 2).
Columna vertebral
Las unidades estructurales en las que se ha fraccionado la columna vertebral siguen una división clásica: cervicales (C), torácicas (T), lumbares (L) y caudales (Ca). La columna vertebral de este ejemplar de rorcual común tiene 58 vértebras estructuradas según la fórmula vertebral: C7 T14 L14 Ca23 (tabla 1, tabla 2) con una longitud total (TCL) de 10,30 m y un peso de 470,95 kg (fig. 2). En la tabla 2 se registran el peso y las medidas de anchura, altura y longitud de cada vértebra junto al análisis comparativo entre ellas.

Tabla 2. Análisis comparativo de la columna vertebral del espécimen Balaenoptera physalus, MZB 83-3084. Table 2. Comparative analysis of the vertebral column of Balaenoptera physalus, MZB 83-3084.
Vértebras cervicales (C)
El cuello está formado por siete vértebras cervicales libres con una longitud del cuerpo que varía desde un mínimo de 4 cm en la C3 hasta un máximo de 10,8 cm en el atlas (tabla 2). El cuello está formado por siete vértebras cervicales libres con una longitud total de 0,47 m y un peso de 29,9 kg (fig. 2). La mayor altura del proceso neural (NPH) corresponde al atlas (24 cm). Las caras anterior y posterior del cuerpo vertebral son ligeramente convexas. Todas las vértebras cervicales presentan metapófisis en la parte anterior de la vértebra y zigapófisis en la parte posterior que coinciden con las metapófisis de la vértebra siguiente, lo que limita el movimiento intervertebral. Las zigapófisis están presentes sólo en las vértebras cervicales y las tres primeras vértebras torácicas. La espina neural del axis (C2) presenta una depresión central en la que se apoya la espina neural del atlas. La relación CL/CH en las cervicales presenta un valor mínimo de 0,20 en C3 siendo este valor el menor para todo el esqueleto vertebral. La relación CH/CW es inferior a 1 (compresión dorsoventral) en todas la vértebras, aunque el atlas se aproxima a la forma circular (0,9) (tabla 2, fig. 3: 1A-1C).

Fig. 3. Análisis gráfico de la columna vertebral del espécimen Balaenoptera physalus, MZB 83–3084. (Para las abreviaturas ver tabla 2.)
Fig. 3. Graphic analysis of the vertebral column of specimen Balaenoptera physalus,MZB 83–3084. (For abbreviations see table 2.)
Las vértebras C3, C5 y C6 tienen el arco neural roto, con pérdida de la espina neural. Las vértebras C3, C4 y C6 han perdido también los arcos dorsoventrales. Estos arcos, aunque incompletos, están inclinados posteriormente hasta la vértebra C5, donde empiezan a inclinarse anteriormente. La vértebra C7 está completa y no presenta arco dorsoventral sino proceso transverso inclinado anteriormente, igual que la espina neural, que es corta (9 cm). En la lámina 2 están fotografiadas las vértebras cervicales.
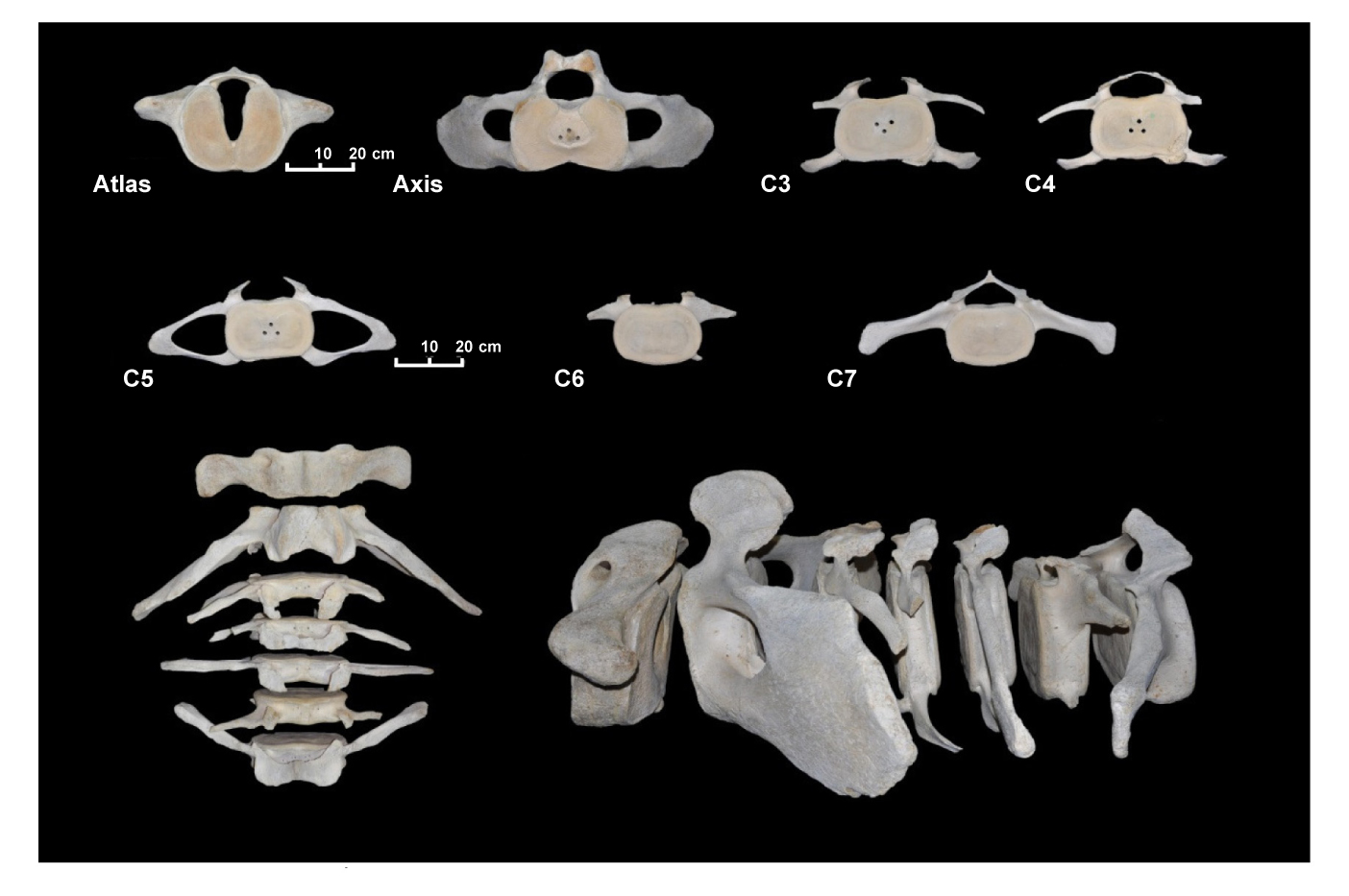
Lámina 2. Vértebras cervicales del espécimen MZB 83–3084 Balaenoptera physalus.
Plate 2. Cervical vertebra of specimen MZB 83–3084 Balaenoptera physalus.
Vértebras torácicas (T)
El tórax presenta 14 vértebras torácicas con una longitud total de 2,57 m y un peso de 139,2 kg (fig. 2). La longitud del cuerpo vertebral en las 14 vértebras torácicas aumenta progresivamente desde un mínimo de 9 cm de la T1 a un máximo de 22 cm de la T14, produciéndose cambios continuos y graduales (tabla 2). El centro de las vértebras torácicas muestra una relación CL/CH en aumento hasta alcanzar la T14 y la relación CH/CW sigue siendo inferior a 1 (entre 0,6 y 0,7), lo que indica que se mantiene la compresión dorsoventral. La altura del proceso neural (NPH) va aumentando desde la T1, con una longitud de 13,7 cm, a la T11, con una longitud de 44 cm (tabla 2, fig. 3: 2A-2C). Desde la vértebra T8 hasta las primeras lumbares, la espina neural presenta deformaciones y fracturas con pérdida de hueso. Los procesos transversos de las vértebras torácicas tienen una inclinación anterior hasta la T9, a partir de la cual empiezan a inclinarse posteriormente. Las metapófisis sobresalen del cuerpo vertebral y encajan con la espina neural de la vértebra anterior, limitando así el movimiento de la columna, sobre todo el movimiento de rotación (lámina 3).

Lámina 3. Vértebras torácicas del espécimen MZB 83–3084 Balaenoptera physalus.
Plate 3. Thoracic vertebra of specimen MZB 83–3084 Balaenoptera physalus.
Vértebras lumbares (L)
Las 14 vértebras lumbares tienen una longitud total de 3,33 m y un peso de 161,7 kg (fig. 2) y presentan una gran uniformidad en la relación CL/CH con una longitud del cuerpo (CL) que varía de 22 cm de L1 a 26,3 cm de L14, lo que supone un rango de 4,3 cm. Este valor es pequeño comparado con el de las vértebras torácicas, en las que el rango es de 13 cm (tabla 2). La relación CH/CW es inferior a 1 (entre 0,7 y 0,8), lo que indica que se mantiene la compresión dorsoventral. La altura del proceso neural de las vértebras lumbares es mayor que en las torácicas y varia de 37,2 cm en la L1 a 44,3 cm en la L14 (tabla 2, fig. 3: 3A-3C). Al igual que las vértebras torácicas, las lumbares presentan deformaciones en la parte superior de la espina que le dan una forma irregular.
La L12 es la primera vértebra en la que se reduce notoriamente la metapófisis, es decir que esta no sobrepasa la parte anterior del cuerpo de la vértebra. La metapófisis de una vértebra ya no se encaja en la espina neural de la vértebra anterior por lo que el movimiento entre vértebras será mayor que en las torácicas. También los procesos transversos se empiezan a reducir notablemente a partir de L5, variando de 37 cm a 25 cm en la L14 (lámina 4).
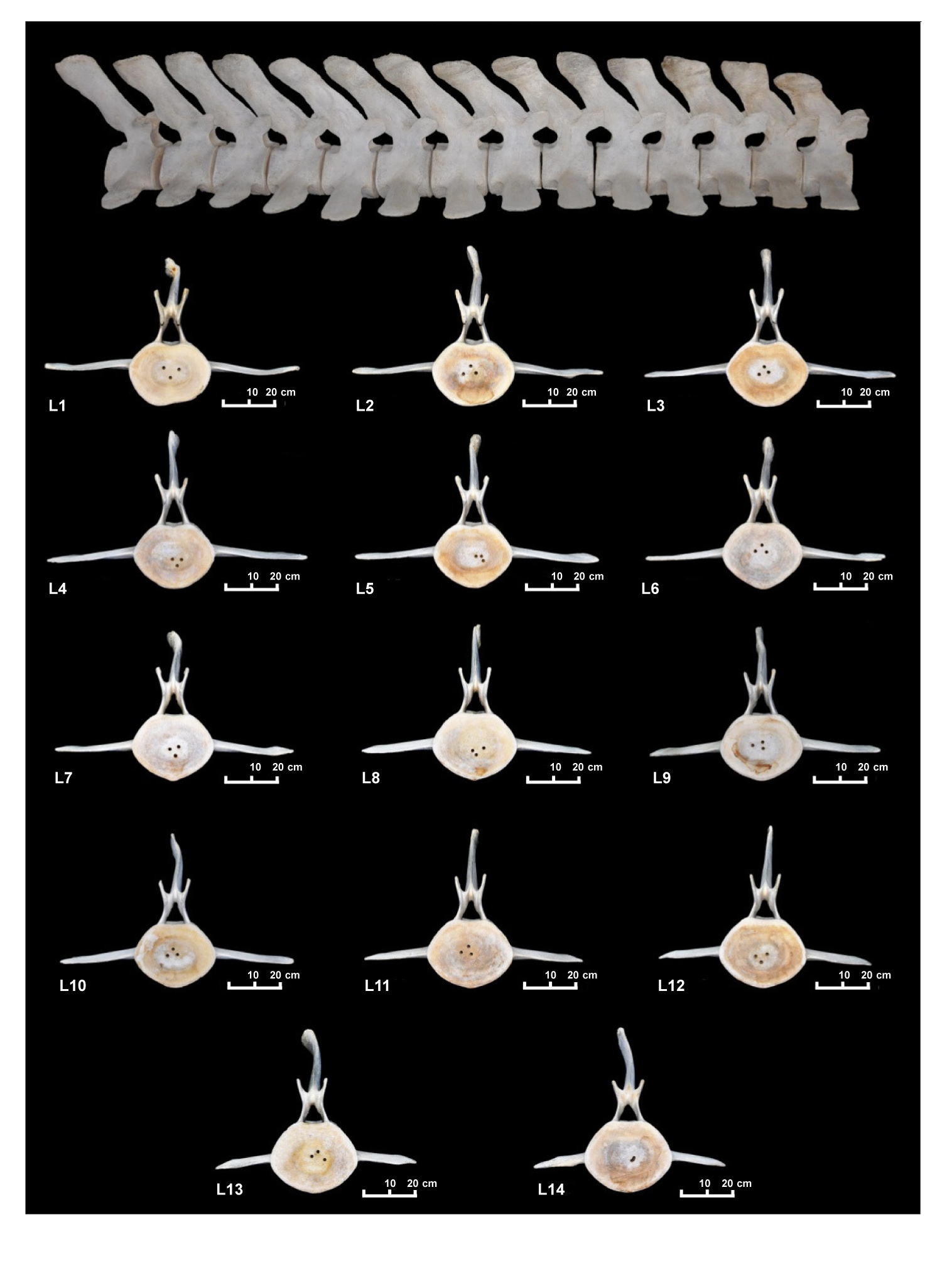
Lamina 4. Vértebras lumbares del espécimen MZB 83–3084 Balaenoptera physalus.
Plate 4. Lumbar vertebra of MZB 83–3084 Balaenoptera physalus.
Vértebras caudales (Ca)
Las 23 vértebras caudales tienen una longitud total de 3,93 m y un peso de 140,15 kg (fig. 2). Las 23 vértebras caudales son las que presentan mayores diferencias morfológicas entre ellas (lámina 5). La longitud del centro varía entre 26,6 cm en la Ca1 a 4,2 cm en la Ca23, lo que supone un rango de variación de 22,4 cm. Los cuerpos vertebrales de las primeras caudales son los que alcanzan la mayor longitud del esqueleto, llegando al máximo en Ca3 con una longitud de 27 cm (tabla 2). Una mayor longitud del centro impide que se produzcan interferencias entre los procesos de vértebras adyacentes y aumenta la rotación intervertebral (Buchholtz & Schur, 2004). Las primeras vértebras caudales presentan compresión dorsoventral, pero a partir de la Ca14 la relación entre CH/CW es mayor a 1 y las vértebras muestran compresión lateral. En la vértebra Ca20 la relación entre la altura y la anchura es de 1,16 (tabla 2, fig. 3: 4A-4C). La vértebra Ca9 es la que presenta mayor anchura (34,5 cm) y mayor altura (29,5 cm) de todo el cuerpo vertebral. A partir de la Ca9 se produce una disminución progresiva del tamaño del centro en sus tres dimensiones (anchura, altura y longitud) llegando a medir la Ca23 5,6 cm de ancho y 4,2 cm de largo. Las vértebras caudales del espécimen se diferencian en dos grupos en términos de longitud relativa del centro. El primer grupo, que corresponde a la zona del pedúnculo caudal, sigue una línea de tendencia negativa, mientras que el segundo grupo, que corresponde a las vértebras de la aleta caudal, sigue una línea de tendencia positiva. La altura del proceso neural va disminuyendo progresivamente hasta la Ca16, en la que el mismo desaparece. En el gráfico 1.4c se observa un punto sinclinal en la vértebra Ca14 que corresponde a una rotura con pérdida de hueso del proceso neural.
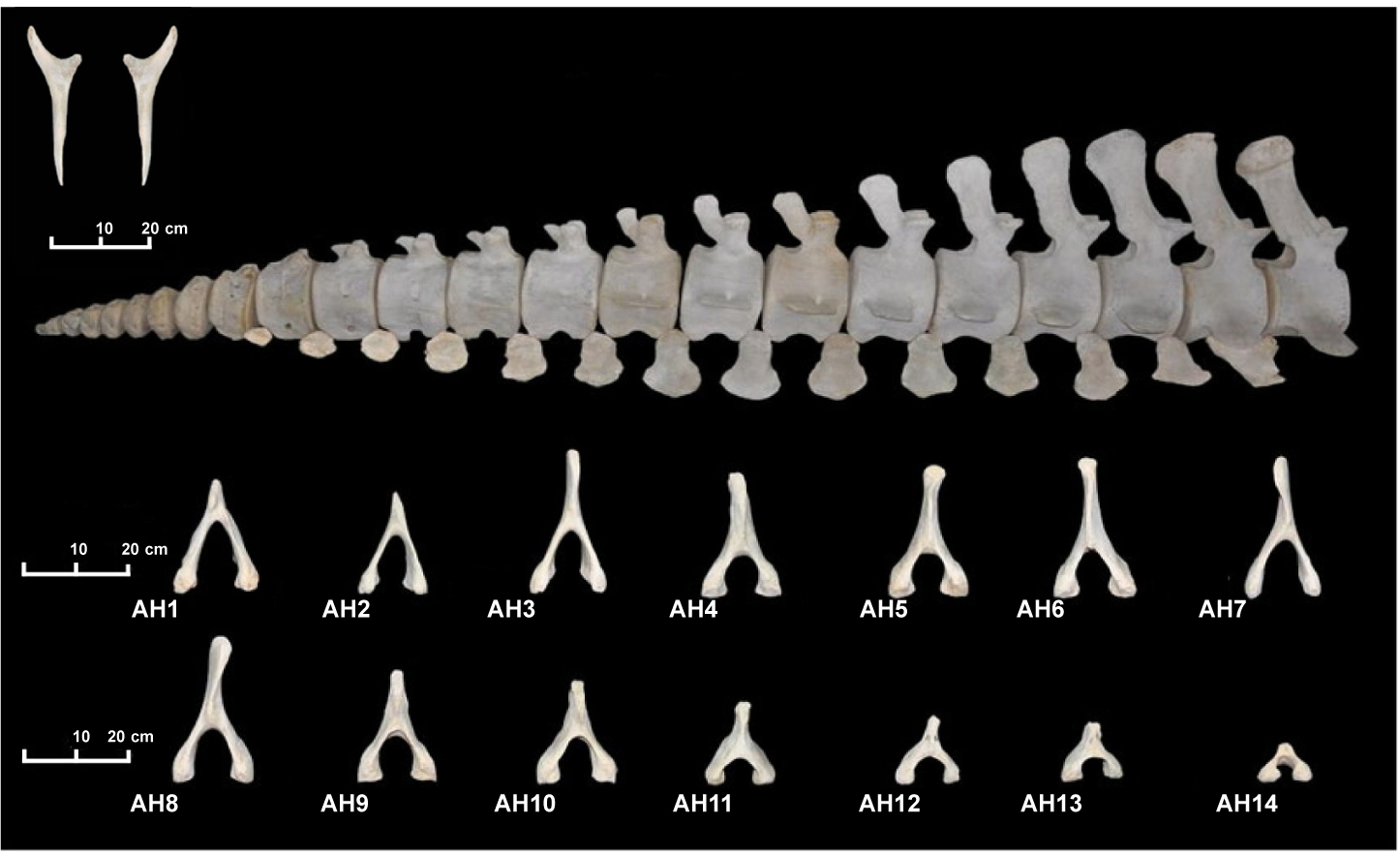
Lámina 5. Vértebras caudales, arcos hemales y huesos vestigiales del espécimen MZB 83–3084 Balaenoptera physalus
Plate 5. Caudal vertebra, hemal arches and vestigial bones of specimen MZB 83–3084 Balaenoptera physalus.
Hemapófisis (arcos hemales)
El espécimen presenta 14 hemapófisis o arcos hemales fusionados en la región caudal. Se articulan entre los bordes de cuerpos vertebrales adyacentes. El primer arco hemal señala la primera vértebra caudal (tabla 1, lámina 5). El peso total de los arcos hemales es de 8 kg (fig. 2).
Costillas
El espécimen presenta 14 pares de costillas (tabla 1), asociadas a las 14 vértebras torácicas, que rodean y protegen las vísceras. También estabilizan la estructura y limitan el desplazamiento y la rotación entre las vértebras. La cabeza de la primera costilla no es doble y su parte distal se ensancha notablemente (19 cm). Las tres primeras costillas presentan un tubérculo que se une a la vértebra por la parte ventral. El ejemplar no presenta costillas esternales. La costilla con menor longitud es la primera, que mide 121,5 cm. La costilla de mayor longitud es la sexta, que mide 210 cm. El último par de costillas presenta una forma completamente diferente al resto, siendo su parte proximal en forma de “s”. El peso total de las costillas es de 13,36 kg (fig. 2). Todas las costillas están fotografiadas en la lámina 6.

Lámina 6. Costillas y esternón del espécimen MZB 83–3084 Balaenoptera physalus.
Plate 6. Ribs and sternum of specimen MZB 83–3084 Balaenoptera physalus.
Esternón
El esternón, situado entre el primer par de costillas, está formado por una única pieza, el manubrio, que en la línea media mide 65,5 cm de ancho y 50,3 cm de largo (tabla 1, lámina 6). El peso del esternón es de 2,5 kg (fig. 2).
Esqueleto apendicular
Huesos vestigiales
Se trata de dos huesos sin unión directa con las vértebras. La homología con los huesos de la pelvis está bien establecida, aunque todavía no está claro con qué hueso de la pelvis son homólogos o si se trata de la fusión de varios. Estos pequeños huesos, que están situados dorsalmente al ano y son anteriores al primer arco hemal, se encuentran inmersos entre el músculo pélvico y tejido conectivo (tabla 1, lámina 5). El hueso vestigial izquierdo que se conserva tiene una longitud máxima de 34,6 cm y un peso de 162 g (fig. 2). El vestigial derecho es una reproducción en madera.
En la tabla 1 se muestran las medidas registradas y en la lámina 7 la osteografía de los huesos que conforman la cintura pectoral.

Lámina 7. Cintura pectoral del espécimen MZB 83–3084 Balaenoptera physalus.
Plate 7. Pectoral girdlewaist of specimen MZB 83–3084 Balaenoptera physalus.
Escápula
La escápula presenta una altura máxima de 92 cm y una longitud máxima de 73,5 cm (tabla 1). El proceso coracoides es alargado, robusto y se extiende anteriormente por la superficie medial alcanzando una longitud máxima de 19,5 cm y una anchura máxima de 9,5 cm. El proceso acromion se sitúa paralelamente al coracoides con una anchura máxima de 12 cm y tiene forma alargada y aplanada por su extremo distal. El peso de cada escápula es de 12,5 kg (fig. 2).
Húmero
La escápula se articula con la cabeza del húmero, que es robusto, corto y está orientado dorsomedialmente. La longitud máxima del húmero es de 48 cm y su anchura máxima en el extremo distal de 27 cm (tabla 1). El peso de cada húmero es de 6,9 kg (fig. 2).
Ulna y radio
El radio tiene una longitud de 77,6 cm y una anchura en su extremo distal de 18 cm (tabla 1). La longitud de la ulna es de 83,5 cm y presenta en la parte proximal un proceso olécranon muy marcado. Las epífisis están totalmente fusionadas, lo que confirma que el ejemplar era un animal adulto y había alcanzado su talla máxima. La ulna y el radio del lado izquierdo pesan 4 kg y 4,5 kg, respectivamente (fig. 2).
Mano
En el extremo distal del radio y la ulna encontramos cinco huesos carpales dispuestos en dos hileras. La línea proximal está formada por tres carpales que, nombrados desde el radio hacia la ulna, reciben los nombres de “radial”, “intermedio” y “lunar”. La anchura transversal de la línea proximal de los carpales mide 28 cm (tabla 1). La línea distal está formada por dos carpales denominados “magnum” y “unciforme”.
La mayoría de las falanges del espécimen eran reproducciones en madera que han sido sustituidas por materiales más duraderos. El dígito I (“pollex”) es el que se sitúa debajo del radio y está compuesto por cinco falanges. El dígito II (“index”) presenta siete falanges. El dígito III (“medium”) es el de mayor longitud y está formado por ocho falanges. El dígito IV (annularis) está formado por cinco falanges. El dígito V no está presente. Los carpales “magnum” y “unciforme”, junto con la falange 1 del digito III (“medium”), se encuentran fusionados en la aleta derecha. En la aleta izquierda, “magnum” y el “unciforme” se encuentran también fusionados (lámina 7). El peso de los carpales y falanges es de 3,3 kg (fig. 2).
Discusión
El espécimen de rorcual MZB 83-3084 Balaenoptera physalus, del Museo de Ciencias Naturales de Barcelona proviene de un varamiento registrado en 1862 en Llançà (Girona). El esqueleto reconstruido con el tejido intervertebral mide 17,35 m de longitud total. El esqueleto completo antes de su montaje tiene una longitud de 14,6 m y un peso de 1.161,59 kg. El sexo no se pudo determinar. En la lámina 8 se pueden observar fotografiados todos los huesos de su esqueleto. Respecto a las diferencias en la talla de la ballena, conviene señalar que la longitud del esqueleto no se corresponde con la longitud que tendría el animal vivo debido a que en las medidas osteológicas no se incluye al tejido fibrocartilaginoso presente entre las vértebras. En nuestro caso, la diferencia entre la longitud conocida a pie de playa (17,35 m) y la del esqueleto limpio (14,61 m) es de 275 cm, lo que equivale al 15,8% menos de lo medido en fresco. En el montaje para determinar la separación entre las vértebras que correspondería al animal vivo hemos tenido en cuenta la longitud medida a pie de playa, por lo que el espécimen montado alcanza una longitud similar a la que tendría la ballena viva. A este respecto, Tomilin (1957) referencia en Holanda (océano Atlántico) un macho de 22 m que pasó a 20,58 m (142 cm; 7,5%) y otro ejemplar en la costa oeste de América del Norte (océano Pacífico) que pasó de 20,74 m a 18,91 (183 cm; 9,9%). Un caso similar se da en un ejemplar varado en 1855 en Cerdeña (Mediterráneo) cuyo tamaño conocido en fresco era de 20 m pero su esqueleto mide 19,3 m. Considerando un 10% de diferencia entre ambas longitudes, el autor del estudio osteológico estima que la ballena mediría unos 21 m (Cagnolaro, 1977).

Lámina 8. Esqueleto del espécimen MZB 83–3084 Balaenoptera physalus: C. Vértebras cervicales; T. Vértebras torácicas; L. Vértebras lumbares; Ca. Vértebras caudales; Ah. Arcos hemales.
Plate 8. Skeleton of specimen MZB 83–3084 Balaenoptera physalus: C. Cervical vertebra; T. Thoracic vertebra; L. Lumbar vertebra; Ca. Caudal vertebra; Ah. Hemal arches.
La longitud del cráneo (4,31 m) representa el 24,84% de la longitud total, lo que coincide con los valores referidos por Aguilar (1995) y con los descritos para otros ejemplares de Balaenoptera physalus conservados en museos de Italia (Nicolosi et al., 1997; Roselli et al., 1997). Según Aguilar (1995), el cráneo de rorcual mediterráneo representa el 20-25% de la longitud total (tabla 3).
Tabla 3. Relación entre longitud total y longitud del cráneo de distintos ejemplares de Balaenoptera physalus. Tabla 3. Relationship between total length and cranial length of several specimens of Balaenoptera physalus.
La longitud del cráneo es cinco veces mayor que la anchura del rostro en su longitud media, valor que coincide con el referido para la especie (Tomilin, 1957).
La columna vertebral tiene 58 vértebras estructuradas según la fórmula C7 T14 L14 Ca23, una longitud total de 10,30 m y un peso de 470,95 kg. A este respecto, Tomilin (1957) señala para el rorcual común un total de 60-63 vértebras, usualmente 62, con la formula vertebral: C7, D15 (16), L14 (13-16), Ca25 (24-27). En la tabla 4 se compara la fórmula vertebral con la de dos ejemplares expuestos en museos italianos. Todas las epífisis de las vértebras están fusionadas y no se aprecian en ellas líneas de sutura, indicando que el espécimen ha alcanzado su talla máxima.

Tabla 4. Comparativa de la fórmula vertebral con ejemplares expuestos en museos italianos. Table 4. Comparison of vertebral formula with specimens exhibited in Italian museums.
Los porcentajes de cada zona (cervical, torácica, lumbar y caudal) respecto a la longitud total de la columna vertebral obtenidos para este rorcual son muy similares a los obtenidos con otros trabajos (tabla 5).
Tabla 5. Porcentajes comparativos de cada zona de la columna vertebral. Tabla 5. Comparative percentages for each segment area of the vertebral column.
La longitud del cuerpo de las vértebras (CL) aumenta gradualmente, a excepción de la C1 (atlas) y la C2 (axis), que tienen una longitud de cuerpo mayor que el resto de las cervicales, hasta alcanzar la máxima longitud en las caudales Ca3 (27 cm) (fig. 4). Desde esta vértebra empieza a disminuir gradualmente hasta la Ca15, donde se inicia una disminución brusca de la longitud hasta la Ca23 (4,2 cm). Una mayor longitud del centro impide que se produzcan interferencias entre los procesos de vértebras adyacentes y aumenta la rotación intervertebral (Buchholtz & Schur, 2004). La altura del cuerpo vertebral (CH) aumenta también gradualmente a partir de C2 llegando a su máxima altura en la Ca7 (29,5 cm) a partir de la cual se produce una continua pérdida de altura (fig. 4). La anchura de las vértebras (CW) mantiene cierto grado de homogeneidad a lo largo de la columna alcanzando la máxima anchura en la L14 y L15 (34,5 cm) a partir de donde se va reduciendo gradualmente hasta la Ca23 (fig. 4).

Fig. 4. Longitud, altura y anchura de los cuerpos vertebrales de la columna.
Fig. 4. Length, height and width of the vertebral bodies of the spine.
La forma del cuerpo de las vértebras en vista anterior presenta una compresión dorsoventral (CW > CH) hasta la Ca14, a partir de la cual la altura empieza a ser mayor que la anchura. Sin embargo, donde se aprecia que la vértebra es más alta que ancha es en la Ca20, justo en la zona posterior del pedúnculo caudal (fig. 5). En las vértebras cervicales, la longitud del centro es muy pequeña y las caras anterior y posterior del cuerpo vertebral ligeramente convexas, lo que provoca alguna interferencia en los márgenes del centro entre vértebras adyacentes con un desplazamiento absoluto mínimo para las cervicales. En la figura 6 se analizan conjuntamente la forma del centro vertebral y la longitud relativa. Se puede observar como la zona con mayor compresión lateral coincide con el punto sinclinal en la longitud relativa del centro. Estos parámetros indican que en esta zona el movimiento de las vértebras es diferente. En la zona anterior, el movimiento es menor, siendo la columna más estable, mientras que a partir del punto sinclinal la columna es más flexible.

Fig. 5. Relación entre la altura y la anchura del cuerpo vertebral.
Fig. 5. Relationship between the height and width of the vertebral bodies.

Fig. 6. Comparación entre la relación altura/anchura y longitud/altura.
Fig. 6. Comparison between the height/width and length/height relationship.
Las vértebras Ca17 y Ca18 forman la base del pedúnculo caudal y a partir de estas empezarían las vértebras que sustentan la aleta caudal.
El proceso transverso alcanza su mayor anchura (TPW) en el axis aunque en el resto de cervicales el proceso se reduce. Las vértebras C3, C4 y C6 presentan los procesos rotos y a partir de C7 los procesos empiezan a aumentar hasta la T14. Desde la T14, los procesos transversales empiezan a reducirse hasta llegar a la Ca13, en la que ya no se hallan presentes. Esta pérdida del proceso transverso se traduce en una mejor rotación sagital de las vértebras (fig. 7). El proceso neural de las vértebras C3, C5 y C6 se encuentra fracturado y, a partir de C7, los procesos neurales van en aumento progresivo hasta llegar a la T7, donde empieza una serie de irregularidades. En las vértebras T8 y T9 se observa un punto de inflexión producido por una rotura con pérdida de hueso de la punta de la espina neural. A partir de la T10, las irregularidades son debidas a deformaciones óseas en las puntas de las espinas neurales de las vértebras. La altura del proceso neural de las vértebras lumbares es mayor que en las torácicas debido a que en las vértebras lumbares y en las primeras caudales se produce una mayor inserción de los músculos destinado a la locomoción. La vértebra Ca2 es la que presenta mayor altura del proceso neural y a partir de esta se produce una disminución progresiva hasta alcanzar la Ca16, a partir de la cual desaparece el proceso neural (fig. 8).

Fig. 7. Longitud del proceso transverso de las vértebras.
Fig. 7. Length of the transverse process of the vertebra.

Fig. 8. Longitud del proceso neural de las vértebras.
Fig. 8. Length of the neural process of the vertebra.
Todas las vértebras, exceptuando las últimas caudales, presentan metapófisis que sirven no solo como anclaje del músculo al proceso neural, sino también como limitación del movimiento de la columna ya que las metapófisis encajan con la espina neural de la vértebra anterior. La L12 es la primera vértebra en la que se reduce notoriamente la metapófisis, es decir que esta no sobrepasa la parte anterior del cuerpo de la vértebra y ya no se encaja en la espina neural de la vértebra anterior por lo que el movimiento de la región lumbar es mayor que el de la región torácica.
En los ejemplares estudiados por otros autores, las costillas más largas son la 6 y la 7 (Tomilin, 1957; Nicolosi et al., 1997), obteniéndose el mismo resultado en el presente trabajo.
El esternón de rorcual tiene muchas variaciones individuales y variaciones relacionadas con la edad. El esternón del rorcual del MCNB presenta una forma parecida al ejemplar adulto procedente de Francia que se reseña en Tomilin (1957). El esternón tiene una forma parecida al de otra ballena del Mediterráneo, la estudiada por Cagnolaro (1977), aunque en ella el esternón no está completamente osificado y presenta un orificio central.
En este artículo se aportan datos detallados de todos los huesos que conforman el esqueleto del rorcual (MZB 83-3084 Balaenoptera physalus) conservado en el MCNB, que muestran grandes similitudes con otros ejemplares adultos estudiados, especialmente del Mediterráneo. Los resultados aquí presentados aportan nuevos datos de la biometría del rorcual común en el Mediterráneo que permitirán realizar comparaciones entre individuos y avanzar en el conocimiento de la estructura ósea y el tipo de movilidad de la especie.
Actualmente, el esqueleto del rorcual se encuentra montado y expuesto en el edificio Fòrum. El MCNB encargó a la empresa GEA un montaje reversible, por lo que en el futuro se podría disponer del esqueleto completamente desarticulado si fuera necesario. El presente trabajo pone a disposición de la comunidad científica los datos de los huesos estudiados individualmente, datos que actualmente no serian medibles sin desmontar el esqueleto.
















