
Arxius de Miscel·lània Zoològica. Volum 17 (2019) Pàgines: 123-144
Coleoptera of Genting Highland, Malaysia: species richness and diversity changes along the elevations
Musthafa, M. M., Abdullah, F.
DOI: https://doi.org/10.32800/amz.2019.17.0123Descarregar
PDFCita
Musthafa, M. M., Abdullah, F., 2019. Coleoptera of Genting Highland, Malaysia: species richness and diversity changes along the elevations. Arxius de Miscel·lània Zoològica, 17: 123-144, DOI: https://doi.org/10.32800/amz.2019.17.0123-
Data de recepció:
- 05/09/2018
-
Data d'acceptació:
- 08/04/2019
-
Data de publicació:
- 08/07/2019
-
Compartir
-


-
Visites
- 1671
-
Descàrregues
- 439
Abstract
Coleoptera of Genting Highland, Malaysia: species richness and diversity changes along the elevations
The objective of this study was to measure beetle richness and diversity in Genting Highland at four major elevations (500 m, 1,000 m, 1,500 m and 1,800 m). Beetles were collected using light traps, malaise traps and pitfall traps. Altogether, 1,499 beetle samples representing 156 morphospecies were collected. Light trap and pitfall traps were more effective than Malaise trap. The 500 m elevation band displayed high species richness, abundance and diversity with all indices showing a decreasing pattern. The species accumulation curve displayed a progressive asymptote for all the altitudinal transects, showing the sampling effort was sufficient for this study. A long-term monitoring program of beetle diversity and distribution is useful to test abiotic factors that might influence biodiversity. This study also serves as a benchmark for further studies on this highly disturbed montane cloud forest in Peninsular Malaysia and will be useful to implement effective conservation management, particularly under the threat of climate change.
Data published through GBIF (Doi: https://doi.org/10.15470/i0uuis).
Key words: Abundance, Beetle, Biodiversity, Ecosystem, Forest
Resumen
Coleópteros de Genting Highland, Malasia: cambios en la riqueza de especies y en la diversidad según la altitud
El objetivo de este estudio es medir la riqueza y diversidad de escarabajos en Genting Highland en cuatro altitudes principales (500 m, 1.000 m, 1.500 m y 1.800 m). Se utilizaron trampas de luz, trampas malasias y trampas de caída (trampas pitfall) para recolectar los escarabajos. En total se recolectaron 1.499 ejemplares de escarabajo correspondientes a 156 morfoespecies con trampas de luz y trampas de caída, que resultaron más efectivas que las trampas malasias. La franja de 500 m de altitud registró la mayor riqueza, abundancia y diversidad de especies, con un patrón decreciente de todos los índices. La curva de acumulación de especies mostró una asíntota progresiva en todos los transectos altitudinales, indicativa de que el esfuerzo de muestreo de este estudio era suficiente. Un programa de monitorización a largo plazo de la diversidad y distribución de escarabajos también sería útil para comprobar los factores abióticos que pueden influir en la biodiversidad. Este estudio servirá asimismo como referencia para posteriores trabajos en esta selva nubosa de montaña considerablemente alterada de la península de Malaca y puede ser útil para implementar una gestión de conservación efectiva, especialmente ante la amenaza del cambio climático.
Datos publicados en GBIF (Doi: https://doi.org/10.15470/i0uuis).
Palabras clave: Abundancia, Escarabajos, Biodiversidad, Ecosistema, Selva
Resum
Coleòpters de Genting Highland, Malàisia: canvis en la riquesa d’espècies i en la diversitat segons l’altitud
L’objectiu d’aquest estudi és mesurar la riquesa i diversitat d’escarabats a Genting Highland en quatre altituds principals (500 m, 1.000 m, 1.500 m i 1.800 m). Es van utilitzar paranys de llum, paranys malaisis i paranys de caiguda (paranys pitfall) per recol·lectar els escarabats. En total es van recol·lectar 1.499 exemplars d’escarabat corresponents a 156 morfoespècies amb paranys de llum i paranys de caiguda, que van resultar més efectius que els paranys malaisis. La franja de 500 m d’altitud va registrar més riquesa, abundància i diversitat d’espècies, amb un patró decreixent de tots els índexs. La corba d’acumulació d’espècies va mostrar una asímptota progressiva en tots els transsectes altitudinals, indicativa que l’esforç de mostreig d’aquest estudi era suficient. Un programa de monitoratge a llarg termini de la diversitat i la distribució d’escarabats també seria útil per comprovar els factors abiòtics que poden influir en la biodiversitat. Aquest estudi també servirà com a referència per a treballs posteriors en aquesta selva nuvolosa de muntanya considerablement alterada de la península de Malacca i pot ser útil per implementar una gestió de conservació efectiva, especialment davant l’amenaça del canvi climàtic.
Dades publicades a GBIF (Doi: https://doi.org/10.15470/i0uuis).
Paraules clau: Abundància, Escarabats, Biodiversitat, Ecosistema, Selva
Introduction
Beetles are good indicators to assess biodiversity in an area, considering biodiversity as species diversity, genetic diversity and ecosystem diversity within an area or biome or biosphere. Higher beetle diversity can generally predict a high diversity of other components of an ecosystem (Cajaiba et al., 2014). Monitoring the beetle diversity can be useful to measure the changes in biodiversity over time (Morrison et al., 2012). Beetles have been widely used in diverse ecological studies due to their favorable characteristics, such as enormous ecological fidelity, high taxonomic diversity, presence in all ecosystems, ease of large-scale collection, and high functional diversity (von Hoermann et al., 2018). Moreover, beetles have been widely studied to assess changes in species diversity along elevational clines worldwide (Tänzler et al., 2015). Even though beetles can be a good model organism for ecological studies, they have been poorly addressed by the scientific community (Escobar et al., 2005; Tänzler et al., 2015). Elevation is one of the key driving forces of biodiversity in montane ecosystems, where anthropogenic activities have higher impacts. Moreover, elevational gradients contribute to the environmental and biodiversity of UNESCO Mountain Biosphere Reserves (MBRs), which offer variety of challenges for conservation under the global climate change (UNESCO, 2006).
Insect distribution pattern along the elevations is contoversial. Though a number of species richness patterns have been suggested, the mostly reported mid-elevational peak comes from short-term sampling regimes with disturbance at lower elevations. Moreover, studies have revealed a long-term sampling strategy tends to produce lower elevational peaks (McCoy, 1990).
Mountains are exciting natural laboratories and they are becoming focal points various ecology and biodiversity related researches. They influence species diversity and created much interest used great among early ecologists (Ficetola et al., 2017). Tropical mountains, moreover, are regarded as hotspots of biodiversity and endemism (Chen et al., 2009; Merckx et al., 2015), and Malaysia is among the biodiversity hotspots in South East Asia. There are number of studies on species diversity patterns along elevational gradients such as land snails (Liew et al., 2010), leaf litter ants (Brühl et al., 1999), litter-dwelling ants (Yusah et al., 2012), black fly (Ya’cob et al., 2016) and butterflies (Ismail et al., 2018; Abdullah and Musthafa, 2019). Diversity studies of beetles have been fairly well covered by the scientific community, but they focused little on the diversity change across elevations in the montane ecosystems. Elevational beetle diversity has not been explored at Genting Highland, Malaysia. Therefore, the objective of this study was to assess and understand species richness, abundance and diversity changes among beetles at Genting Highland along the elevations. We used multiple trapping methods and compared compositional differences (beta diversity) between elevations. Moreover, we compared three different types of trapping methods
Material and methods
Sampling sites
The mountains in Malaysia, the Titiwangsa Range, are located in the centre from Pahang to Kelantan states. Genting Highland is on the Pahang and Selangor border in Bentong District, which is just 50 km from Kuala Lumpur, Malaysia.
Genting Highland is the most disturbed area. The entire summit region has been replaced by amusement parks, casinos and hotels (Peh et al., 2011). Before the conversion of Genting Highland into an entertainment site, this area was a virgin, undisturbed forest that could be reached only via jungle trekking (Stone, 1981; Piggott, 1977). The flora of Genting Highland characterizes a sparsely distributed upper montane cloud forest and elfin forest in Peninsular Malaysia (Stone, 1981). Plant diversity at Genting Highland includes more than 460 species of flowering plants and around 100 species of ferns and fern allies, with, 28 of these flowering plants being endemic and three species considered rare. Loss of biodiversity at Genting Highland is highlighted by few studies. Hasanah et al. (2009) reported a loss of 81 % of the Pteridophytes recorded by Piggott (1977), while 47 orchid species were threatened according to Anggerik et al. (2012). Chua and Saw (2001) also discussed the drastic drop in floral diversity at Genting highland with immense environment changes.
Faunistic diversity of Genting Highland includes around 18 amphibian, 134 bird, 42 mammal and 18 reptile species. Genting Highland accommodates 20 % mammal, 21 % bird, 8 % reptile and 21 % amphibian species among the total of described animal species in Peninsular Malaysia. Moreover, Genting Highland is home to 51 % of known highland mammal species, 50 % of bird species, 34.6 % of reptile species and 60 % of the amphibian species in Peninsular Malaysia (WWF Malaysia, 2002). From the IUCN Red List of threatened species (2017), two species of mammals, serow (Capricornis sumatrensis) and Indochinese tiger (Panthera tigris corbetti), are recorded as endangered. Aetholops alecto, a grey pygmy fruit bat, a strictly montane forest species and Lygosoma miodactylum (Single finger larut skink) are rare species that are also found in Genting Highland, and Hystrix brachyuran (Malayan porcupine) has been reported as vulnerable according to the IUCN (2017).
Experimental design and beetle sampling methods
Beetles were collected from sites at 500 m, 1000 m, 1,500 m and 1,800 m a.s.l. (fig. 1). Light traps, Malaise traps and pitfall traps were used to collect the beetles. Two light traps, two Malaise traps and 25 pitfall traps with five sets of pitfall traps arranged in a diagonal shape were fixed at each elevation. Non-baited wet pitfall traps were 200 ml plastic cups (65 mm diameter, 9.5 cm depth) filled with 50 ml 70 % alcohol, sunk into the ground with the brim at ground level and large leaves were positioned at the same level to protect the traps from flooding . Malaise traps were made of nylon net with a collection jar half filled with 70 % alcohol. They were fixed to the branch of a tree not more than 1.5 m from the ground and they were also fixed for 24 hours. Light traps were made of mosquito netting with a 160 watt mercury bulb connected to a portable Honda EU10i portable power generator. It was fixed just above ground level and beetles attracted to the light were collected using collection bottles. Light traps were fixed for six hours at each elevational band from 6 p.m. to 12 a.m. midnight. Sampling was replicated four times in 2015/2016 (December 2015, March 2016, August 2016 and November 2016)

Fig. 1. Sampling location altitudes within Genting Highland in Peninsular Malaysia. Fig. 1. Altitud de los puntos de muestreo en Genting Highland, península de Malaca.
Specimen identification and tallying
All the collected samples were sorted and tallied to morphospecies level using established keys (Triplehorn and Johnson, 2005) and then cross checked with the Wildlife Department of Malaysia, University of Malaya, National University of Malaysia and Forestry department of Malaysia museum collections. The previous collection at our lab was also used to identify samples to morphospecies level.
Statistical analysis
The commonly used nonparametric estimators ACE (Abundance-based Coverage Estimator), ICE (Incidence-based Coverage Estimator) and Chao 1estimators were used to calculate species richness at each altitudinal band using PAST 3.07 (Hammer et al., 2001). The Clench model was used to estimate the sampling effort efficacy with the use of estimated species. Richness and slope of the species accumulation curve for all beetles collected from each elevational transect were plotted using Statistica 8.0 (StatSoft Inc., 2007).
Species abundance was calculated for all sampling methods at altitudinal band and differences between these values were analyzed with Kruskal-Wallis nonparametric tests. For diversity analysis, the Shannon diversity index, the Simpson diversity index and Fisher’s alpha diversity indexes were used, while the Margalef index was used to calculate calculation. Cluster analysis for abundance was conducted in Statistica 8.0, using a dissimilarity matrix with the Bray-Curtis index as a distance measure, and the Ward´s amalgamation algorithm. Beta diversity was measured through Bray-Curtis index of similarity as for the faunistic similarity between the four altitudinal sites. Cluster analysis was also performed, using PAST 3.07 (Hammer et al., 2001) to define groups of sites according to species composition, using the Bray-Curtis index as a distance measure and the UPGMA (Unweighted Pair-Groups Method using arithmetic Averages) method as an amalgamation algorithm.
Results
The sampling effort resulting from the species accumulation curve displayed progressively increasing curves for four altitudinal bands (fig. 2) and did not fully reach the asymptote; this is generally expected in the tropics due to the high specious nature of diverse beetles (Escobar et al., 2005; Chao et al., 2009). From four altitudes (table 1), 156 different beetle species were collected, representing 35 families, 98 genera and 1,560 specimens (table 2, GBIF dataset: https://doi.org/10.15470/i0uuis). Almost one third of the total collected specimens were singletons (53 out of 156 species), while 17 species were doubletons. Singletons refer to only one specimen collected, whereas doubletons mean two specimens. Families dominating the overall fauna were Staphylinidae (14 species 297 individuals, 20.78%), Scarabaeidae (14 species, 264 individuals, 18.59 %), Bostrichidae (eight species, 202 individuals, 14.23 %), Chrysomelidae (17 species, 132 individuals, 9.30 %) and Carabidae (13 species, 108 individuals, 7.61 %). The most abundant species was Crypturgus sp. 1 representing 5.63 % of the total number of individuals (80 individuals), followed by Lymantor sp. 1 (4.37 %, 62 individuals), while the next five species were very closely followed by each other in numbers, Apogonia sp. 1, Apogonia sp. 4 (3.24 %, 46 individuals), Aleocharinae sp. 1 (3.17 %, 45 individuals), Stenus sp. 1 and Stenus sp. 2 (3.03 %, 43 individuals). The highest number of specimens, species, genera and species were obtained from lower elevation (500 m), whereas the pattern showed a liner decrease along the gradients. Table 2 lists the collected species based on elevations.

Fig. 2. Species accumulation curve of beetle species collected at Genting Highland from four elevations sampled. Fig. 2. Curva de acumulación de especies de las especies de escarabajos recolectadas en las cuatro altitudes de muestreo de Genting Highland.

Table 1. Number of individuals, species, genus and families collected from the four altitudinal bands at Genting Highland. Tabla 1. Número de ejemplares, especies y familias recolectados en las cuatro franjas de altitud de Genting Highland.

Table 2. Species collected from four elevational gradients (E, in m) at Genting Highland: MT, Malaise trap; PT, pitfall trap; LT, light trap. Tabla 2. Especies recolectadas en los cuatro gradientes de altitud (E, en m) de Genting Highland: MT, trampa malasia; PT, trampa de caída (pitfall); LT, trampa de luz.
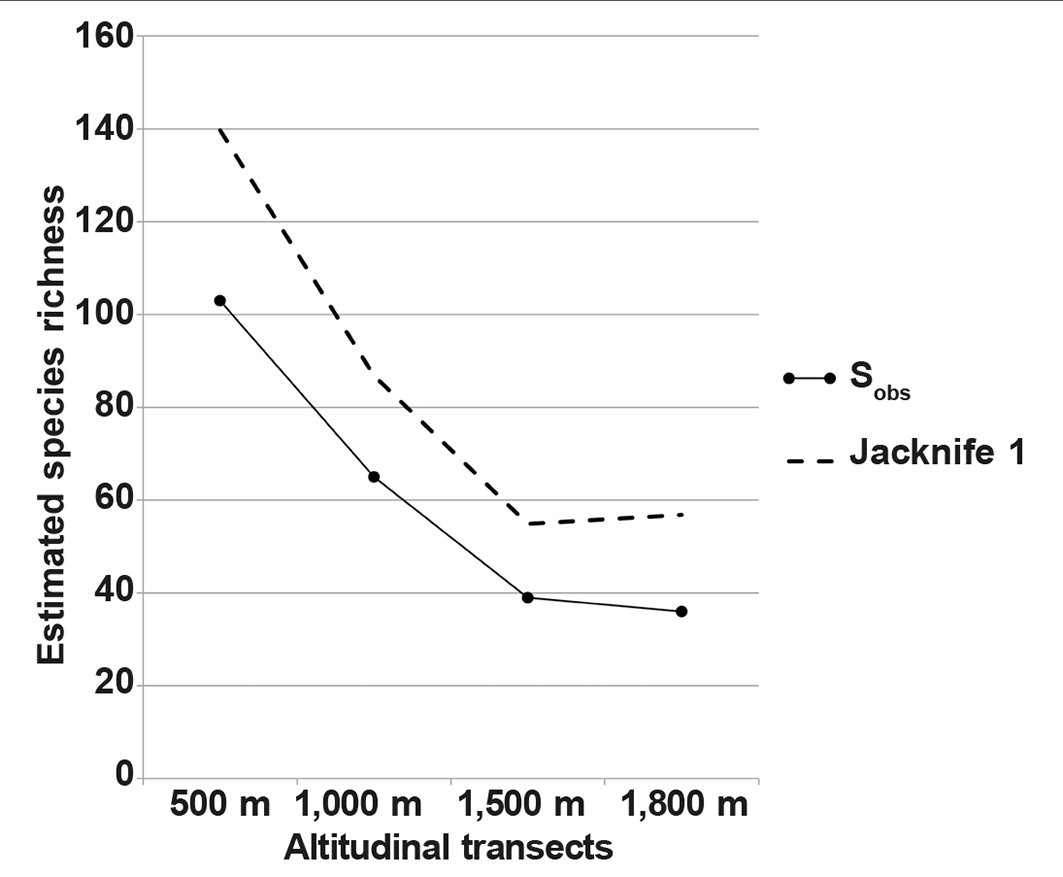
Table 3 shows the species richness measures using the observed, estimated and Clench model results; the observed number of species richness differs significantly between elevations. Species richness was highest at 500 m elevation where the nonparametric estimates (ACE, ICE and Jackknife 1) showed highest values and the Clench model also showed highest species richness at 500 m. These estimates of species richness showed highest values at 500 m altitude (ACE = 137.79; ICE = 135.51; Jackknife 1 = 161.83; Clench model = 108.63). The estimated richness decreases linearly when moving along the elevations. The slope values for Clench model assessment showed values that were slightly higher than the cut-off value of 0.10 for 500 m, 1,000 m and 1,800 m. Observed species richness (Sobs) values, species diversity and abundance values showed a decreasing pattern with an increase in altitude at Genting Highland (fig. 3).

Table 3. Observed (Sobs) and estimated (Sest) species richness of beetles from Genting Highland Malaysia, based on nonparametric indices and the Clench model. Tabla 3. Riqueza de especies de escarabajos observada (Sobs) y estimada (Sest) en Genting Highland, Malasia, basada en índices no paramétricos y en el modelo de Clench.
Fig. 3. Estimated species richness (Sobs and Jackknife 1) pattern across the elevational transects observed at Genting Highland: Sobs, observed species richness. Fig. 3. Patrón de riqueza de especies estimada (Sobs and Jackknife 1) en los transectos altitudinales observados en Genting Highland: Sobs, riqueza de especies observada
A comparison of the observed species richness (Sobs) across the trapping methods showed a higher value for 500 m light trap (Sobs = 52), 500 m Malaise trap (Sobs = 41) and 500 m pitfall trap (Sobs = 38) (table 4). Nonparametric estimates (ACE, ICE and Jackknife 1) also displayed high values for 500 m light trap. The clench model slope value showed higher values around 0.1, which clearly indicated the underestimated values for light traps. Overall collection from pitfall resulted in a high number of specimens collected (n = 588; 41.41 %), followed by the light trap (n = 444; 31.27 %) and the Malaise trap (n = 388; 27.33 %).

Table 4. Shannon diversity index, Simpson diversity index, Fisher alpha diversity index, and Margalef abundance indice for four altitudes from Genting Highland, Malaysia. Tabla 4. Índice de diversidad de Shannon, índice de diversidad de Simpson, índice de diversidad alfa de Fisher e índice de abundancia de Margalef en cuatro altitudes diferentes de Genting Highland, Malasia.
Highest values for diversity indices (Shannon diversity index, Simpson diversity index and Fisher alpha diversity index) and abundance index (Margalef index) were displayed at 500 m elevation of Genting Highland (table 4). The values for diversity measures decreased linearly when moving up the elevations, where the lowest values for all indices were found at 1,800 m except for the Fisher alpha diversity index which showed lowest value at 1,500 m. The Shannon diversity index was higher than 3.0 for all four elevations while Simpson diversity index was above 0.9, indicating that Genting Highland has high beetle diversity. The cluster of abundance of species present according to the elevations resulted in three distinctive groups (fig. 4). According to the species abundance, 1,500 m and 1,800 m elevations formed two different faunistic groups, while 1,000 m and 500 m formed the third faunistic group. Moreover, cluster analysis for beta diversity resulted in three distinguishable groups on the basis of the difference in beetle composition between elevation sites: group 1-500 m and 1,000 m clients; group 2-1,500 m and group 3-1,800 m (fig. 5).

Fig. 4. Cluster analysis for species abundance at the four tested elevations (gray dotted line indicates the delimitation of groups). Fig. 4. Análisis clúster de abundancia de especies en las cuatro altitudes estudiadas (la línea de puntos gris indica la delimitación de grupos).

Fig. 5. Cluster analysis grouping different elevational sites in Genting Highland, calculated via Bray–Curtis dissimilarity index using the UPGMA method. Fig. 5. Análisis clúster agrupando emplazamientos a diferentes alturas de Genting Highland, calculado mediante el índice de disimilitud de Bray–Curtis y empleando el método UPGMA.
Discussion
Species richness is possibly the basic measure of biodiversity and it is vital to assess biodiversity in any ecosystem (Gotelli and Colwell, 2011). Tropical ecosystems of earth harbor mega-diverse arthropod communities and this study showed a good number of beetle species richness from Genting Highland, the most disturbed montane cloud forest in Malaysia. Moreover, this is the first and most inclusive study of beetle fauna from different altitudes using three different trapping methods.
The three types of traps appeared to impact the capture of different beetles in relation to abundance, richness, and their attractiveness. The species composition differed highly between the traps at elevation, reiterating the importance of using multiple trapping methods for beetle sampling, especially in the tropics. Furthermore, the sampling methods complemented each other. More intense sampling strategies should be attempted to capture other beetle taxa from other localities as well. The contrasting trap resulted from this study also supports the use of multiple sampling techniques targeting different beetle taxa (Basset et al., 2007), reiterating the point that there is no silver bullet for tropical arthropod sampling strategies.
Assessing the diversity of invertebrate species from tropical regions can be expected to be incomplete and biased regarding sampling region, habitat and taxon (Novotny, 2007) and hard to achieve in any part of the world (Chao and Chiu, 2016). Species’ accumulation curves are therefore useful to assess the sampling effort using the level of asymptote of the curve. Theoretically, reaching the asymptote means sampling until no more new species are found, but practically, this cannot be achieved even with an extended biodiversity assessment study (Longino et al., 2002). From this study, the species accumulation curve for elevations showed an ever increasing curve and did not reach the asymptote from all transects, mainly due to the presence of a good number of singletons and doubletons from the sampling sites. Conversely, richness estimators imply that the number of species would be greater if researchers used other complementary sampling approaches that would increase the number of species found and reduce the number of singletons and doubletons. Therefore, the overall sampling efforts from Genting Highland beetles from four altitudes were within the acceptable limits, and further extensive sampling strategies might result in a higher beetle catch. In future studies, if the target taxon is ground dwelling, it will be advisable to use pitfall traps only, while if the focus is on flying insects, both light traps and Malaise traps can be used.
The use of these species richness estimators in the form of ACE, ICE and Jackknife 1 are recommended in other studies (Colwell et al., 2012). Of these estimators, Melo and Froehlich (2001), Rico et al. (2005) and Basualdo (2011) concluded ICE and ACE would be the best fit for a better general performance across these estimators. This study also reiterated the use of multiple species richness estimators because different estimators consider different statistical models. Therefore, this study proposes the non-parametric estimators such as ACE, ICE and Chao 1 were excellent tools both to estimate species richness and to measure completeness of inventories of some of the most conspicuous groups of beetles.
The most popular metric to quantify biodiversity composition is the Shannon diversity index (Nagendra, 2002). Margalef (1972) notes that these values are typically between 1.5 and 3.5 and rarely exceed a value of 4. Based on this scale, it can be concluded that the beetle diversity in the studied locality is high (Shannon > 3.00), where at 500 m altitudinal transect the Shannon diversity index even exceeded 4.00. The Simpson diversity index also showed higher values (> 0.93) for all the altitudinal clines, whereas the Fisher alpha diversity values were also above 10. Fisher’s alpha diversity measure is regarded as one of the most useful and often recommended indicators of community diversity due to its independence from sample size (Beck and Schwanghart, 2010). Caution has been advised with the use of the Shannon and Simpson diversity indices to assess the biodiversity since they are highly associated with situations and sites, and thus require greater importance during interpretation (Nagendra, 2002; Morris et al., 2014). Therefore, there is no single diversity measure that can be applied to all situations as a universal parameter (Morris et al., 2014).
A decrease in the species richness pattern has been observed in some studies (Linzmeier and Ribeiro-Costa, 2009). The present study also showed a decreasing pattern of species diversity at Genting Highalnd, Malaysia. The climatic factors, evolutionary drivers and land use pattern play a prominent role in shaping the biodiversity of montane ecosystems (Merckx et al., 2015). As suggested by McCoy (1990), the sampling strategies also play a part in species richness patterns in montane ecosystems. Moreover, there might be other reasons. The composition of a plant community around the locality (Silva and Hernandez, 2014) might impact of climatic factors (Polesel and Damborsky, 2017) and thermal adaptability of species. These factors could regulate the distribution and local adaptation of beetles over elevational gradients, since temperature intensely influences their flight activity (García-Robledo et al., 2016). Furthermore, according to Kubota et al. (2000), good numbers of beetle species have a relatively low dispersal ability compared with other insects and are thus vulnerable to isolation by geographic barriers.
The information presented here provides baseline data that allows for comparisons of the diversity and species richness of beetles on a regional and national scale. This information could be used as an initial step to analyze the potential use of beetles as a bioindicator group in Malaysia. To gain further understanding, future studies on different beetle communities in Malaysia need to be oriented towards patterns of specific species and their links with environmental variation, and also towards interplay between ecosystem components and beetle species. Environmental influences on species diversity are vital to implement effective conservation management, particularly under the effects of rapid climate change.
Acknowledgements
This study was financed by Vot RP004E/13SUS and PG059/2014B. We would like to thank Mr. Uriel Jeshua Sánchez-Reyes, Mr. Mohd Shukri and Mr. Davindram A/L Rajendram.

















{kind=link}