
Arxius de Miscel·lània Zoològica. Volumen 18 (2020) Páginas: 123-142
Structure of waterbird assemblages in fragmented coastal wetlands of Northeastern Algeria
DOI: https://doi.org/10.32800/amz.2020.18.0123Descargar
PDFCita
Bouldjedri, M., Mayache, B., 2020. Structure of waterbird assemblages in fragmented coastal wetlands of Northeastern Algeria. Arxius de Miscel·lània Zoològica, 18: 123-142, DOI: https://doi.org/10.32800/amz.2020.18.0123-
Fecha de recepción:
- 11/11/2019
-
Fecha de aceptación:
- 18/09/2020
-
Fecha de publicación:
- 05/10/2020
-
Compartir
-


-
Visitas
- 1458
-
Descargas
- 322
Abstract
Structure of waterbird assemblages in fragmented coastal wetlands of Northeastern Algeria
This study was carried out from 2007-2017 at the ecological complex of the Jijel wetlands in the north-east of Algeria. Censuses were conducted during the twelve months of each year to study variation in richness and abundance of waterbird populations for each season. We used distance sampling (point count and transect methods). A total of sixty species (eleven orders and sixteen families) were recorded. The Anatidae and Scolopacidae families were the most numerous with thirteen species. The common coot (Fulica atra), and the Mallard (Anas platyrhynchos) were the most abundant and frequent species each year. About 70 % of the species recorded occur as migrants, passing between the western Palearctic and their winter quarters in North Africa. Phenologically, we found 15 % were breeders, and from the point of view conservation status, 56 % were rare, 40 % were protected by Algerian regulations and 8 % were threatened species listed in the Red List of the International Union for Conservation of Nature (IUCN). Our results of ornithological monitoring in the wetlands in Algeria show that action is needed to address the consequences between birds, human activities, and climate change.
Data published through GBIF (Doi:10.15470/oud0fp)
Key words: Breeding, Eco-complex, Habitat, Waterbirds, Wetlands, Wintering
Resumen
Estructura de las poblaciones de aves acuáticas de los humedales fragmentados de la costa del noreste de Argelia
Este estudio se realizó durante el periodo 2007-2017 en el complejo ecológico de los humedales de Jijel, en el noreste de Argelia. Se efectuaron censos durante los doce meses de cada año para estudiar las variaciones de riqueza y abundancia de las poblaciones de aves acuáticas en cada estación. Utilizamos el muestreo a distancia (punto de recuento y método de transectos). Registramos un total de 60 especies (de 11 órdenes y 16 familias). Las familias Anatidae y Scolopacidae fueron las más numerosas, con 13 especies. La focha común (Fulica atra) y el ánade azulón (Anas platyrhynchos) fueron las especies más abundantes y frecuentes cada año. Aproximadamente el 70 % de las especies registradas son migrantes de paso entre el Paleártico occidental y sus zonas de invernada en el norte de África. Desde el punto de vista fenológico, encontramos que el 15 % son reproductoras. Por lo que respecta a su estatus de conservación, el 56 % son raras, el 40 % están protegidas por las regulaciones argelinas y el 8 % son especies amenazadas incluidas en la Lista Roja de la Unión Internacional para la Conservación de la Naturaleza (UICN). Este seguimiento ornitológico sugiere que los humedales de Argelia requieren un proceso que tenga en consideración y armonice la interferencia entre aves acuáticas y actividad humana, así como el cambio climático global y sus consecuencias como, por ejemplo, la sequía.
Datos publicados en GBIF (Doi:10.15470/oud0fp)
Palabras clave: Cría, Eco complejo, Hábitat, Aves acuáticas, Humedales, Invernada
Resum
Estructura de les poblacions d’ocells aquàtics dels aiguamolls fragmentats de la costa del nord-est d’Algèria
Aquest estudi es va portar a terme durant el període 2007-2017 al complex ecològic dels aiguamolls de Jijel, al nord-est d’Algèria. Es van fer censos durant els dotze mesos de cada any per estudiar les variacions de riquesa i abundància de les poblacions d’ocells aquàtics en cada estació. Vam utilitzar el mostreig a distància (punt de recompte i mètode de transsectes). Vam registrar un total de 60 espècies (d’11 ordres i 16 famílies). Les famílies Anatidae i Scolopacidae van ser les més nombroses, amb 13 espècies. La fotja comuna (Fulica atra) i l’ànec collverd (Anas platyrhynchos) van ser les espècies més abundants i freqüents cada any. Aproximadament el 70 % de les espècies registrades són migrants de pas entre el Paleàrtic occidental i les seves zones d’hivernada al nord d’Àfrica. Des del punt de vista fenològic, vam trobar que el 15 % són reproductores. Pel que fa a l’estatus de conservació, el 56 % són estranyes, el 40 % estan protegides per les regulacions algerianes i el 8 % són espècies amenaçades incloses a la Llista Vermella de la Unió Internacional per a la Conservació de la Natura (UICN). Aquest seguiment ornitològic suggereix que els aiguamolls d’Algèria requereixen un procés que tingui en consideració i harmonitzi la interferència entre ocells aquàtics i activitat humana, així com el canvi climàtic global i les seves conseqüències com, per exemple, la sequera.
Dades publicades a GBIF (Doi:10.15470/oud0fp)
Paraules clau: Cria, Ecocomplex, Hàbitat, Ocells aquàtics, Aiguamolls, Hivernada
Introduction
Wetland habitats are used by waterbird species for nesting, breeding, feeding, sheltering, migration stopovers, and wintering in various stages of their annual life cycle. These ecosystems have high biodiversity and are sites of conservation concern due to the extensive food chain (Saygili et al., 2011). The great diversity of wetlands in North Africa has long been recognized (Gauthier-Lièvre, 1931), but despite concerns for their conservation, they continue to be disturbed, drained and polluted (Bouldjedri et al., 2011; Daoud-Bouattour et al., 2011; Rhazi et al., 2012).
According to the General Directorate of Forestry, Algeria has 2,375 wetlands. They are composed of 2,056 natural areas and 319 human-made wetlands. Although most of these freshwater hydrosystems (lakes, lagoons, marshes, wet meadows, peat lands and alder swamps) are concentrated in the northeastern part of the country, several shallow lakes, with varying salinity are found in the High Tell and the Algerian arid and Saharan zones (‘Chotts’, ‘Sebkhas’, ‘Gueltas’ and Oases). These ecosystems are part of a large Mediterranean eco-complex which is strongly exploited by many waterbird populations, often structured as metapopulations (Boulkhssaim et al., 2006; Samraoui et al., 2011). Several studies show that these zones meet one or more of the criteria required for an Important Bird Area ‘IBA’ (Samraoui and Samraoui, 2008; Baaziz et al., 2011; Benradia et al., 2018; Nouidjem et al., 2019) and constitute a valuable network of sites used each year by thousands of birds migrating between their breeding grounds in the Arctic and their wintering sites in southern Europe and North Africa.
The Jijel region in Northeast Algeria occupies a privileged location for waterbird migration because of its eco-complex of coastal wetlands and geographical situation in the Mediterranean flyway. It therefore plays a vital role as a migratory stopover for aquatic avifauna when they cross the Mediterranean Sea. The few preliminary studies (De Bélair and Samraoui, 2000; Mayache, 2008) performed to date for the area have illustrated the ornithological and ecological value of these wetlands; this ecoregions host notably endangered and vulnerable species registered in the IUCN Red List of threatened species (IUCN, 2019), such as the ferruginous duck (Aythya nyroca), the white-headed duck (Oxyura leucocephala) and the common pochard (Aythya ferina).
Despite their ecological interest, this wetland eco-complex is under strong anthropogenic disturbances, including agriculture, grazing, hunting and fishing (Bouldjedri et al., 2011). This worrying situation motivated the present study, which was carried out over ten years. It describes for the first time the communities of waterbirds that use this area of the Mediterranean flyway and discusses the potential utility of this area as a staging, resting and wintering place. The objectives of the study were to: a) characterize the waterbird communities that visit these coastal wetlands; b) analyze their temporal evolution; c) assess the diversity of each communities and establish comparisons between them; d) assess the importance of these wetlands to propose, where appropriate, protective measures; and e) to complete data on distribution and breeding aquatic birds so as to develop action plans for species threatened by human pressure on Algerian wetlands.
Material and methods
Study area
Fieldwork was conducted in Jijel eco–complex wetlands located in north-eastern of Algeria (36º 34′-36º 52′ N, 5º 33′-6º 19′ E), which are approximately 60 km2 in size. This natural wetland reserve comprises two major aquatic habitats that may vary in vegetation composition and structure, namely: Beni-Belaid Lake and El-Kennar Marsh (see map in fig. 1). This latter freshwater swamp is dominated by herbaceous rather than woody plant species. These mosaic hydrosystems are characterized by several types, mainly lakes, marshes, reservoirs, ponds, estuarine waters, and shallow seawater. Together, they form a complex ecosystem of wetlands. It is of note that the extent of many wetlands declined significantly over the ten–year study period, and that the difference in surface area between the two main sites and their associated wetlands is large, with more than 50 % being less than 3 ha (pers. obs.).
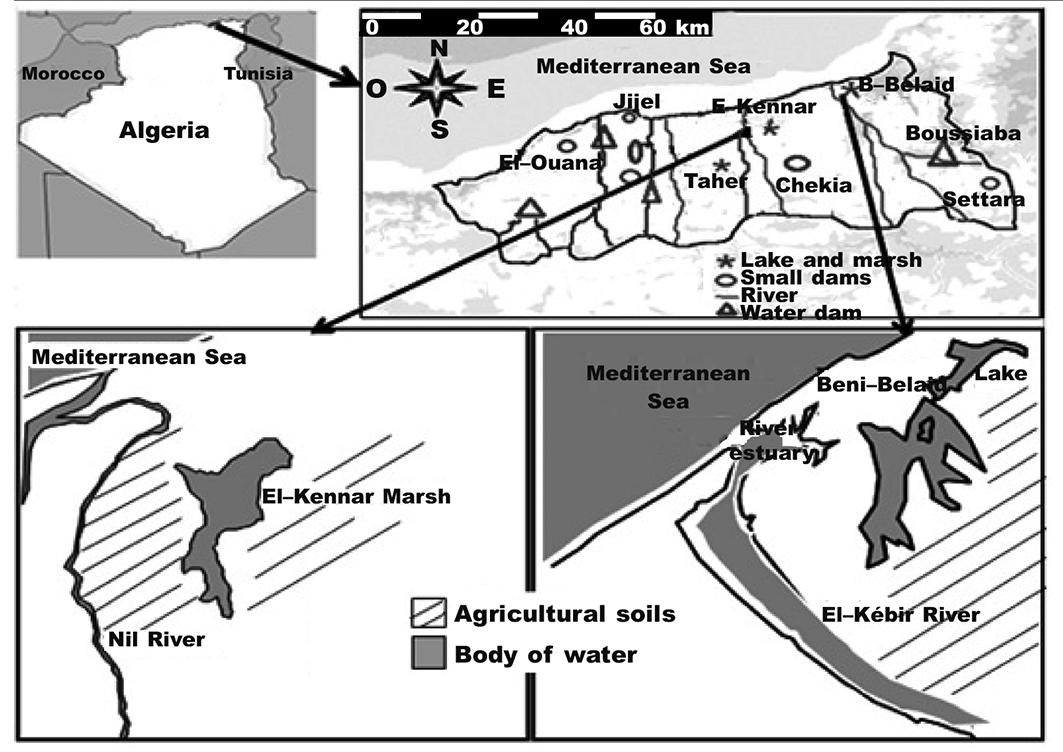
Fig. 1. Map of Jijel wetland eco-complex, north-east Algeria, showing the two main sites: Beni-Belaid Lake and El-Kennar Marsh and their nearby sites. Fig. 1. Mapa del complejo ecológico de humedales de Jijel, en el noreste de Argelia, que muestra los dos puntos de muestreo principales: el lago Beni-Belaid y el pantano El-Kannar y los puntos cercanos a los mismos.
This ecoregion has a typical Mediterranean climate characterized by alternating wet and dry seasons, with a mean annual rainfall of 950 mm, mainly from November to February (Bouldjedri, 2013). The water level peaks between November–April, and drops to its lowest level over August-October.
Beni-Belaid Lake is situated in the east of the mouth of the El-Kebir River. It covers an area of about 43 ha and has brackish and alkaline water. This lake was formed by an abandoned channel of the El-Kebir River and is defined as an old delta of this River (Bouldjedri et al., 2011). During the lowest rainfall period, the water storage depth of the lake often maintains a level of some 0.5-1 m in the centre. The hydrological gradient also has a temporal component, characterized by an intra-annual cyclicity linked to the summer development of the hydrophilic and halophytic communities. The lush vegetation of this site is characterized by woody and herbaceous species, which is structured in belts. It is composed mainly of white poplar (Populus alba), French tamarisk (Tamarix gallica), white broom (Retama raetam), sea clubrush (Bolbochoenus maritimes), and common clubrush (Schoenoplectus lacustris), as well as fields of grass (Cynodan dactylon) and knotgrass (Paspalum distichum). Lake communities are dominated by fennel-leaved pondweed (Potamogeton pectinatus), water horn wart (Ceratophylum demersum), stone warts
(Charophyta), and the invasive species creeping water primrose (Ludwigia peploides). In the context of preservation of the avian biodiversity, Beni-Belaid Lake is listed as a natural reserve and a wetland of ‘international importance’ by the Ramsar Convention (2003).
El-Kennar Marsh is a large open water area. The depth of water storage does not exceed 0.9 m in the centre of the swamp during the drought period. It is well-known that this swamp is contaminated by urban sewage, and it is also under pressure because of intensive illegal hunting. It is an unprotected freshwater swamp of 15 ha with an eutrophic waterbody, characterized by an extensive bed of floating vegetation of European white waterlily (Nymphaea alba) and emergent vegetation such as sea clubrush, common reed (Phragmites australis), common clubrush, branched bur-reed (Sparganium erectum), and Southern cattail (Typha domingensis). A small stand of tamarisk trees is found on its eastern shore. This natural environment is a suburb of the Jijel coastal town that has undergone demographic growth over the last two decades. Since 1997, the site has been surrounded by burgeoning housing and farms. Both Beni-Belaid Lake and El-Kennar Marsh are affected by the same human activities (e.g. agriculture, hunting, grazing, and fishing).
Field survey
To investigate the spatial and temporal variations of waterbird communities, we carried out monthly surveys over ten years, from October 2007 to December 2017, at the two main sites and their related areas. Surveys were carried out during the migration season (in autumn, from September to November), the wintering period (from December to February), the breeding season (from March to July), and the post-nuptial period (from June to August). The species and the peak in total number recorded over a month were all used for analysis. Birds were identified and counted using either: 7 × 50 binoculars or 20 × 60 Meopta telescope, and field guidebooks; we selected waterbirds from the avian fauna that are obligate users of the wetland. Other species were all excluded from the analysis. At each site, all birds were counted, so that the time effort was proportional to the size of the wetland. For each main site, we simultaneously counted its related sites to avoid errors due to bird movements between adjacent areas, the observation points were always the same. Methods of bird observation are numerous and dependent on the studied species and the purpose of the study. Depending on the circumstances, we combined two main methods in our field survey, the absolute method and the relative method.
In the case of the absolute method the census is called exhaustive because the population was estimated directly on its absolute value and all the individuals were counted (point counts). An individual bird count was carried out when the group contained less than 200 birds and when it was at a distance of less than 200 m. This is because the area of the related sites was small enough for us to observe the whole site from one location with no optical obstacle, and one point counting was used (Gregory et al., 2004).
Concerning the relative method, the population was estimated using sampling that includes only a part of the population. We estimated a sample of average size, then divided the field of vision into several transects (i.e. the flock size was divided into small equal blocks) and counted the number of blocks. This method is commonly used in winter counts of waterfowl when the individual number exceeds 200 birds or when estimating at a distance of more than 200 m, as described by the ‘Scan Sampling Method’ (Lamotte and Bourlière, 1969; Blondel, 1975).
For the transect survey, the perpendicular searching distances varied depending on the shape of the site, with 0.5 and 0.8 km in El-Kennar Marsh and Beni-Belaid Lake, respectively (i.e. depending on the size, and boundary location of the site). The vegetation type (emergent, submerged, grasses, shrubs, and trees) was determined by direct observation within the consistency of the point count stations. We also estimated the depth of the water in the shallow area from the length of the legs of shorebird species. Variability in observer error was minimized by measurements being performed by the same observer wherever possible throughout the study. Table 1 lists all the species included in the analyses.
Data analysis
The study of the avifauna ecology was approached by determining ecological indexes directly linked to the balance of populations (Shannon-Weaver, Frequency and Uniformity). The monthly mean number for species richness and the total number of individuals was calculated for the whole period. These values were then used to calculate the diversity of species for the two main sites, expressed by the alpha diversity (Hα) of Shannon-Weaver index (H′): Shannon-Weaver index: it was computed using the following mathematical formula (Shannon-Weaver, 1949):
H‘ = – pi log2 pi
where pi = (ni/N), N is the total population (individuals of all species), ni is the size of the population of species and S denotes the total number of species composing a population (specific richness).
Frequency index: Relative abundance (RA %) is computed by using the formula:
RA (%) = ni / N × 100
where ni is the number of individuals of species taken into account and N is the total number of individuals of all species. Species relative abundance status (table 1) was estimated from the frequencies of sightings over 12 months and the categories were assigned according to the Pettingill (1969) scale: abundant (90-100 %), common (65-89 %), moderately common (31-64 %), uncommon (10-30 %) and rare (1-9 %).
Uniformity Index (Pielou index): this index measures the degree of numerical balance between species represented by the Equitability index (Evenness index), E is calculated as the ratio:
E = H′obs / H′max
between observed diversity H′obs and maximum diversity H′max = log2(S), S being the total number of species recorded during each month (Pielou, 1966). The equitability index makes it possible to assess the imbalances that the diversity index cannot detect; it varies from 0 to 1. When E < 0.5, this means that the individuals mostly tend to belong to a single species. When E > 0.5, it means that species have similar or balanced abundance, the same abundance occurs when the value is exactly one (Okpiliya, 2012).
Statistical analyses
For the entire study region, the mean (± standard deviation) was calculated for each ecological index (Shannon-Weaver diversity (H′), Equitability index (E), Abundance, Specific richness and Monthly population dynamics) based on all observations. The results were compared using analysis of variance (ANOVA), the choice of a parametric test analysis (two ways ANOVA) is supported by the Shapiro-Wilk test on the ANOVA residuals for the previous ecological index comparison which finds no indication that normality is violated. After this analysis, we used a Bartlett test on the ANOVA residual because the normality and homoscedasticity conditions were met: as the p-value was largely greater than 0.05, the assumption of homogeneity of the residual variances was therefore accepted. However, ANOVA led to the conclusion that there were differences between the means in some groups. Tukey’s (HSD) multiple comparison tests were used to compare the difference between each pair of monthly means. The choice of a non-parametric test (Friedman rank-sum test) analysis in the monthly abundance of shorebirds population is referred to that the condition of residual normality is violated (p = 0.001). These tests were performed using the statistical software R version 3·2·2 (R Core Team, 2015).
Correspondence analysis (CA) was applied (bi-plot species and dates) to the data collected from the entire e eco-complex. CA is a two-dimensional solution that is well-suited to describe the spatial and temporal gradients in composition occurrence, status and assembly of waterfowl found at this ecological site. This last statistical analysis was carried out using the ADE-4 software package.
Results
Abundance and species composition
The study shows that Beni-Belaid Lake and El-Kennar Marsh, as well as their nearby freshwater bodies, were frequented by many waterbirds during the wintering and breeding seasons (3,100 and 999 individuals, respectively). In late autumn and at the beginning of winter, progressive arrivals were recorded. The monthly variation of total numbers from the two main sites showed a Gaussian pace (fig. 2A). The temporal distribution of the populations culminating in December for Beni-Belaid Lake (1,250 birds), and in January for El-Kennar Marsh (1,850 birds), all species combined (fig. 2A).
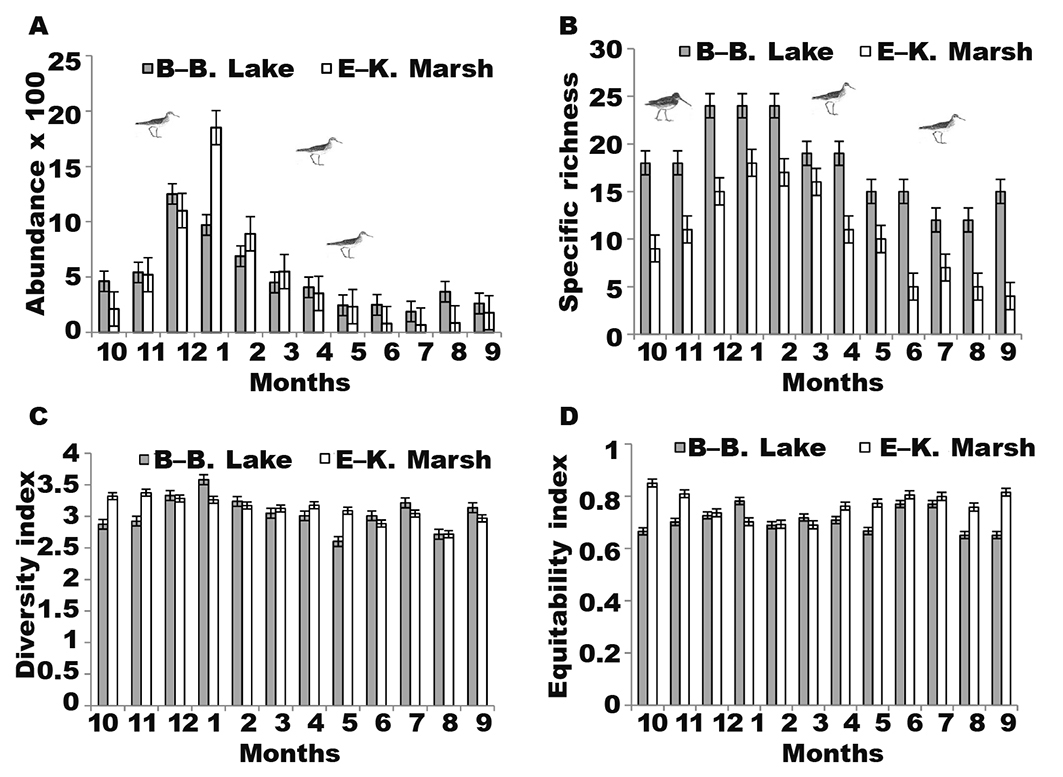
Fig. 2. Seasonal (monthly) variation of waterbird abundance (A), species richness (B), Diversity Shannon–Weaver Index (C), and Equitability Index (D) (± SD) in Beni–Belaid Lake (October 2007–December 2017) and El–Kennar Marsh (June 2007–December 2017). Fig. 2. Variación estacional (mensual) de abundancia de aves acuáticas (A), riqueza de especies (B), índice de diversidad de Shannon–Weaver (C) e índice de equidad (D) (± SD) en el lago Beni–Belaid (octubre de 2007–diciembre de 2017) y en el pantano El–Kennar (junio de 2007–diciembre de 2017).
The bird population of all the sites consisted of 60 species (table 1; datapaper published through GBIF: Doi:10.15470/oud0fp) belonging 16 families and 11 orders. These 60 species represent more than 15 % of Algerian avifauna, with a cumulative population of 12,254 individual birds (appendix 1). About 8 % of the species (n = 5) were listed in the IUCN Red List 2019 and 40 % (n = 24) are listed as nationally protected species (JORADP, 2012). Two families have the same highest number of species: Scolopacidae and Anatidae (13 spp. = 21 %). Ardeidae was represented by seven species (12 %), with the cattle egret (Bubulcus ibis) being the most abundant species. The Rallidae family was represented by four species, among which we found three resident breeders: (1) the common coot (Fulica atra), which is much less secretive than most of the rail family; (2) the common moorhen (Gallinula chloropus); and (3) the purple swamp hen, which exhibited an increase in breeding pair number (three to four) in the two main sites. Accipitridae, Alcedinidae, Burhinidae, Ciconiidae, Haematopodidae, Phalacrocoracidae and Phoenicopteridae were represented by a single species only. Table 1 shows important species richness and the presence of 34 rare taxa at a regional scale. Twenty-six were considered common with season maxima regularly exceeding 300 individuals (appendix 2). However, the ANOVA table showed significant differences in bird abundance between winter and spring, and between summer and autumn (p < 0.05). Tukey test (HSD) analysis showed the only significant differences between months: Dec-Apr, Jan-Apr, May-Jan, Jul-Dec, June-Dec, Jul-Jan, Jun-Jan, at p < 0.01 and May-Dec, Jan-Apr, Jan-Aug, Dec-Aug, Jan-Aug, at p < 0.05.

Table 1. Composition of waterbird assemblage’s occurring at Jijel wetlands eco-complex during the exploration period (2007–2017), with the codes and ecological status, two scales are used: degree of commonality or rarity according to seasons and degree of threat (IUCN category): * the 60 species recorded in our two main wetlands (Beni-Belaid Lake, El-Kennar Marsh) and their annexed sites. Information on rarity and commonness at the regional scale (RC) is given according to Pettingill (1969). Status: PS, protected species, given according to (JORADP, 2012); B, breeding; R, resident; S, summering; V, visitor; W, wintering migrant were given according to our monitoring. , IUCN category: EN, endangered; LC, least concern; NT, near threatened; VU, vulnerable. Tabla 1. Composición de las poblaciones de aves acuáticas del ecosistema de los humedales de Jijel durante el período de exploración (2007–2017), con los códigos y el estado ecológico. Se utilizan dos escalas: grado de frecuencia o rareza según las estaciones y grado de amenaza (categoría de la UICN): * las 60 especies registradas en los dos humedales principales (el lago Beni-Belaid, el pantano El-Kennar) y zonas anexas de acuerdo con (Pettingill, 1969). Se proporciona información sobre rareza y frecuencia a escala regional (RC) según Pettingill (1969). Estatus: PS, especies protegidas, se clasifican de acuerdo con (JORADP, 2012); B, reproductor; R, residente; S, estival; V, visitante; W, migrante invernal según nuestro monitoreo. Categoría de la IUCN: EN, en peligro de extinción; LC, menor preocupación; NT, casi amenazado; VU, vulnerable.

Appendix 1. Number of waterbirds species observed during each season in each wetland category. Apéndice 1. Número de especies de aves acuáticas observadas durante cada temporada en cada tipo de humedal.

Appendix 2. Mean number of birds observed for each species and for each season in both wetlands. Apéndice 2. Número medio de aves observadas para cada especie y cada temporada en ambos humedales.
Diversity patterns
In Beni-Belaid, the diversity index was highest in the winter season (January) with H′ = 3.581, as compared to the lowest value (H′ = 2,601) recorded in spring (May) (fig. 2C). For the second site, El-Kennar Marsh, the highest value of H′ was in late autumn season (November) with H′ = 3.37, while the lowest value (H′ = 2.71) was recorded in summer (August). From the ANOVA table, we can conclude that the difference between sites and monthly diversity was not statistically significant (p > 0.05).
The values of the Pielou index (E) are high for both sites every month (fig. 2D). E > 0.5 means that abundance was similar for all species (Okpiliya, 2012), the maximum being obtained in winter (January) with E = 0.781, and the minimum in early autumn (E = 0.651) (fig. 2D). From the ANOVA table, only the variability between sites showed a significant difference (p < 0.05).
From a ‘specific richness’ aspect, our study area was more frequented during winter, where for both sites a maximum of 42 species was recorded during January (fig. 2B), thus equaling the ornithological value of the most important wetlands in Algeria (Samraoui et al., 2011; Metallaoui et al., 2014; Benradia et al., 2018; Nouidjem et al., 2019).
The significant difference for waterbird specific richness between the two habitats (Beni-Belaid Lake and El-Kennar Marsh) was compared using the two-way analysis of variance (ANOVA) and Tukey’s (HSD) test. The ANOVA table showed that the difference for both sites and months was statistically significant (p < 0.001).
According to their wintering status, Anatidae and Rallidae started to settle in the two sites in late August in relatively low numbers (fig. 3A).Their number increased gradually to reach a maximum of 505 individuals in December Beni-Belaid Lake and 975 individuals during January in El-Kennar Marsh. Large concentrations of ferruginous duck, common moorhen (Gallinula chloropus), common coot, and purple swamp hen, were noted in the relatively unobstructed water beaches at the two main sites not near the disturbance (human activity and poaching). We compared the significant difference for Anatidae and Rallidae abundance between the two habitats was compared by applying the two-way analysis of variance (ANOVA) and Tukey’s (HSD) test. The ANOVA test indicated no significance between the two sites; the output of the Tukey test showed the only significance was between months in the summer and winter (p < 0.05).
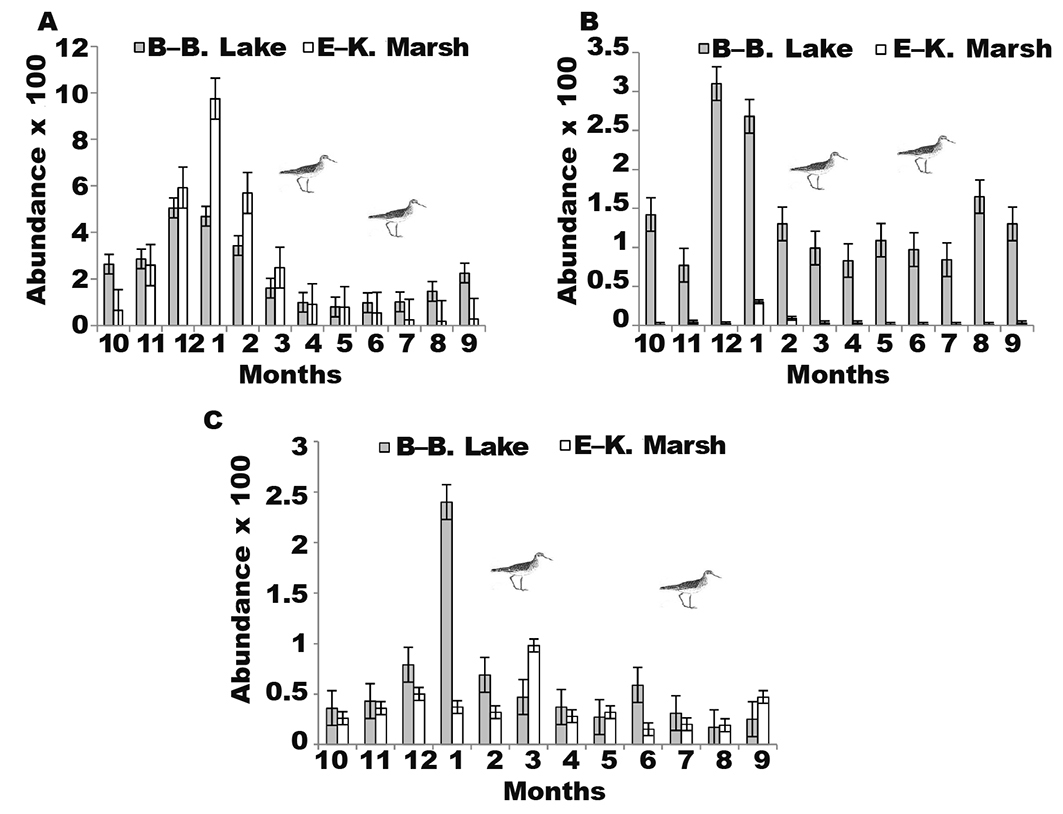
Fig. 3. Monthly dynamics of the population of the main families and groups (variations in abundance ± SD): Anatidae and Rallidae (A); waders and Laridae (B); shorebirds (C), in Beni–Belaid Lake (October 2007–December 2017) and El–Kennar Marsh (June 2007–December 2017). Fig. 3. Dinámica mensual de población de las principales familias y grupos (variaciones en el número total ± SD): Anatidae y Rallidae (A); limícolas y Laridae (B); aves costeras (C), en el lago Beni–Belaid (octubre de 2007–diciembre de 2017) y en el pantano El–Kennar (junio de 2007–diciembre de 2017).
Because of its location near the Mediterranean Sea and the river estuary, Beni-Belaid Lake is often frequented by all kinds of species groups (appendix 2): wading birds, composed mostly of cattle egrets, little egret, purple heron (Ardea purpurea), grey heron (A. cinerea), and white stork; swimming birds (waterfowl, grebes, cormorants Phalacrocorax carbo); and the cosmopolitan seabird yellow-legged gull (Larus michahellis). These observations are in concordance with observations of Güitrón–López et al. (2018) during the winter over five years at Sayula Lagoon, Jalisco (Mexico). These groups of birds use Beni-Belaid Lake as a hiding and refuge area in December-January, and relatively high numbers were observed (310 individuals) (fig. 3B). The group of shorebirds reached its peak (240 individuals) in January (fig. 3C). In El-Kennar Marsh, the peak number recorded was 98 individuals for shorebirds, mainly observed in March. From the Friedman rank sum test table, we can conclude that differences in sites and months were not statistically significant (p > 0.05).
Discussion
Although the ornithological monitoring in the present study covers a limited area, the surveyed avifauna represent more than half of the waterbird species described in Algerian wetlands (97 species identified by Samraoui et al., 2011). This biodiversity assessment indicates that this hydrosystem has retained regional importance for the resident and migrant species of aquatic avifauna since Beni-Belaid Lake was designated a Ramsar site in 2003 (De Bélair and Samraoui, 2000). In this regard, we can say the ecological interest of this eco-complex is comparable to that in the great emblematic wetlands of North Africa, the case of (1) the wetlands of Merja-Zerka in Morocco (Qninba et al., 2009; Cherkaoui et al., 2017), (2) the wetland complex of ‘Eastern Numidia’ in Algeria (Stevenson et al., 1988; Samraoui and De Bélair, 1997), (3) and the wetlands of ‘Kroumiria’ in Tunisia (Daoud-Bouattour et al., 2011; Nefla and Nouira, 2016).
The discussed area should be seen as a network of habitats integrated within the broader network of natural and artificial wetlands represented by the small dams of Chekfa, El-Aouana, Settara, and Tleta; the great dams of Boussiaba, El-Agrem and Kessir, and the swamp of Redjla-Taher (fig. 1). These wetlands can be divided into three major wetland systems: (i) Lacustrine, (ii) Palustrine and (iii) Estuarine, with their heterogeneous vegetation types and habitat (wet grassland, meadows with Juncus, dune and riparion zones) originating from complex water level dynamics. These different hydro systems merit classification as an important international migratory corridor and stopover habitat for aquatic avifauna in Northeast Algeria. Sixty bird species (migratory and resident) are ecologically dependent on only one habitat for at least one part of their annual cycle. It is already recognized as an important area for purple swamphen (Porphyrio porphyrio), Eurasian thick-knee (Burhinus oedicnemus) and the near-threatened ferruginous duck, which use the site for wintering and breeding (De Bélair and Samraoui, 2000). However, our investigation shows that the lacustrine system (the case of Beni-Belaid Lake) is the preferred refuge for most species during all seasons of the year (table 2). We also note the presence in this habitat of the white-headed duck, which is globally threatened due to the significant reduction in its populations over the last decade (Gürsoy-Ergen, 2019; IUCN, 2019).

Table 2. Specific richness of waterbirds occupying the various habitats of the Jijel wetlands eco–complex. Tabla 2. Riqueza específica de aves acuáticas que ocupan los diferentes hábitats del ecocomplejo del humedal de Jijel.
Several studies have been performed in northeastern Algeria but the sampling effort was generally weaker than ours, both spatially and temporally. These previous studies reported temporal variation in the numbers of waterfowl according to the seasons of one or two annual cycles. Metallaoui and Houhamdi (2010) reported 47 species belonging to 15 families during one wintering period (August-April); Boudraa et al. (2014) recorded 53 species (15 families) over one year; Elafri et al. (2016) described 52 species belonging to 13 families over two annual cycles; and Nouidjem et al. (2019) reported 36 species, 11 families in the southeast of Algeria. From the point of view of diversity pattern, all these ornithological studies showed that species belonging to the Anatidae family were widely distributed and occurred in all kinds of habitats (open water zone, shore zone, vegetation zone and riparian area) and the gathering and abundance of numerous duck species and coots were conditioned by the diversity of the existing vegetation cover. These findings corroborate our field observations.
Results in the ornithological literature show that the distribution of waterfowl and waders is strictly linked to specific water regimes, ranging from major flooding to total drought (Pyrovetsi and Papastergiadou, 1992; Catsadorakis et al., 1996). This indicates that water level changes influence the physical structure of habitats, the presence of safe breeding sites, and food accessibility (Rajpar and Zakaria, 2011; Güitrón-López et al., 2018). Therefore, the interplay of several factors, such as the presence of core refuges, landscape structures and water depth, affects the richness and abundance of aquatic avifauna. In this context, Hattori and Mae (2001) highlight that species richness and density of waterbirds is highest in reed beds of aquatic vegetation where the water level is 20-65 cm in depth. Behney (2020) specifies that dabbling ducks can generally only feed at depths of up to 40 cm.
The results of our ornithological study show a spatiotemporal variation in bird assemblage. The migration peak of wintering migrants of shorebirds and ducks, which are two largest groups in our study areas, is from early October to late December. The first waterbirds that arrive at the beginning of the wintering season tend to occupy optimal sites, so as the winter progresses, the best sites are saturated. Later migrant individuals tend to occupy marginal sites. Early April to mid-May is the migration peak of late passage migrants, such as whiskered tern (Chlidonias hybrida), black-winged stilt (Himantopus himantopus), ruff, spotted redshank, and common redshank. These late migrant birds tend to occupy the related habitats of the two main sites (Beni-Belaid Lake and El-Kennar Marsh).
From late spring to autumn, most of the bird population is concentrated in Beni-Belaid Lake, while from late autumn to early winter El-Kennar Marsh receives the highest concentration of birds. These communities are mainly composed of groups of breeding birds. The cattle egrets nest in large trees of the riparian zones, and the dabbling and diving ducks (mallard, ferruginous duck), little egret (Egretta garzetta), purple swamp hen, common moorhen and common coot are often seen in the dense stands of emergent vegetation on the margins and open water; both these habitats are highly appreciated by waterfowl, especially Anatidae and Rallidae (Pirot et al., 1984; Saygili et al., 2011). Many of them nest on floating vegetation and some in riparian vegetation. We add that these birds usually disperse on the shallower lakeshores and muddy areas of the swamp, which are rich in amphibians and invertebrates, an important component of their diet (Kasprzykowski et al., 2014; Metallaoui et al., 2014). It should also be noted that waterbirds are known to adapt their food requirements to prey availability in their habitat (Metallaoui et al., 2014;
Navedo et al., 2017); prey availability varies according to meteorological conditions, availability of trophic resources, and water depth (Saygili et al., 2011; Pérez-Crespo et al., 2013; Datta, 2014; Kasprzykowski et al., 2014; Bouriach et al., 2015; Fouzari et al., 2015). These structural dynamics clearly show nested assemblage models for waterbird communities and may be important determinants of aquatic system evolution.
In term of diversity pattern, our ornithological study shows that species richness reaches its maximum at both sites during the wintering season (December-February). We recorded 24 species at Beni-Belaid Lake and 18 at El-Kennar Marsh (fig. 2B). In North Africa, this period corresponds to the pre- and post-nuptial gathering and transit periods for many waterbirds (Boudraa et al., 2014). Concerning the two indexes relating to the population balance (Shannon-Weaver Diversity Index and Uniformity Index), maximums were noted during the winter season, and mainly during the postnuptial transit period, when the individual representativeness of the species occupying the sites is practically the same, in other words, when the species are represented in more or less equal numbers. These indexes obtained during our study are analogous to those recorded in similar coastal wetlands of North-eastern Algeria during the wintering period (H′ = 3.5 and E = 0.7) by Metallaoui and Houhamdi (2010) and Elafri et al. (2016) at Garaet Hadj-Tahar and Lake Tonga, respectively. Furthermore, the two main sites are adjacent to the Mediterranean Sea and are especially important stopovers for long-distance migratory bird species, including spoonbill (Platalea leucorodia), greater flamingo (Phoenicopterus roseus), and Eurasian curlew (Numenius arquata), which explains the highest level of species richness and diversity notably in winte particularly at Beni-Belaid Lake. The fact that this ‘Ramsar’ site is more attractive for shorebirds can be explained by the large extent of feeding habitats suited to this group of birds, such as mudflats/sandpits and estuarine wetlands. After the wintering season, waterbird abundance and species richness reached their lowest values (12 species at Beni-Belaid Lake and 7 at El-Kennar Marsh). This is due to the departure of all wintering and staging migrant birds to their breeding sites in the Palearctic and sub–Saharan Africa (Samraoui et al., 2011), leaving only nesting birds (sedentary nesters and summer breeders) in the body of water.
Temporal and phenological groupings of avian communities
Despite evident disturbance activities in the area, especially changes in land use and pollution by discharges from municipal wastes, these sites are an important wetland for various functional groups of waterbirds, as shown by the many migratory and resident species recorded there with different phenological status: wintering species accounted for 39 % of all the observed birds, followed by visitors and resident breeders species in the same proportion (25 %), migratory breeders (5 %), and summering species (6 %). It is of note that we have noticed that small groups of some species (great egret Ardea alba and Eurasian spoonbill Platalea leucorodia, black-winged stilt, and white stork Ciconia ciconia) have begun to spend the summer in our coastal wetlands.
The factorial map of CA also allows us to observe a true temporal distribution of the wetlands occupation during the annual cycle. The rate of inertia was: 0.50, 0.15, 0.09, 0.06, 0.05 and 0.04. The first dimension accounted for 50 % of the total variance, while the second accounted for 15 %. The plan created by factors 1 and 2 therefore summarizes 65 % of the total inertia (fig. 4). The first axis showed a clear opposition between summering and wintering populations, demonstrating the succession of temporal occupation of this eco-complex by water birds. However, three populations were discriminated by CA: wintering populations, summering and visitor populations, and breeding populations. Wintering populations consisted mainly of: dabbling ducks (Northern pintail (Anas acuta), Northern shoveler, gadwall, Eurasian wigeon and common teal); diving ducks (common pochard, ferruginous duck, tufted duck and scaup, grebes (great-crested grebe and black-necked grebe (Podiceps nigricollis), great cormorant); and waders (dunlin (Calidris alpina), the common snipe (Gallinago gallinago) and Northern lapwings (Vanellus vanellus). Summering and visitor (passing) species consisted mainly of the spoonbill, knights (spotted redshank: Tringa erythropus, common redshank: Tringa totanus), ruff sandpiper and greater flamingos. These visitors are vagrants from Europe and Asia. Breeding populations were mainly represented by species of the families Anatidae, Ardeidae and Rallidae, and little ringed plover (Charadrius dubius). We observed several behavioural patterns that indicated the breeding status, such as carrying nest-building material and particular aggression (active territorial defense). We also had the opportunity to observe the little ringed plover nest on sandy soil in the open area, with both males and females taking turns to incubate the eggs.
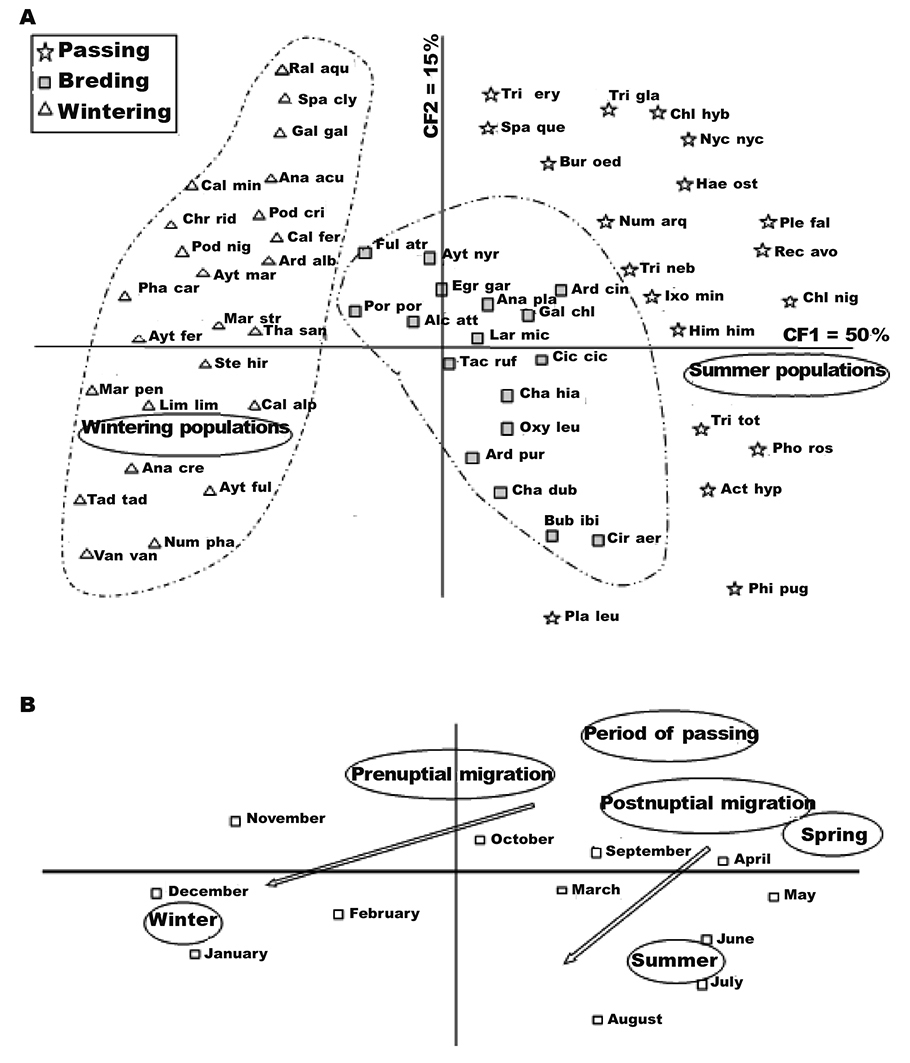
Fig. 4. Ordination plot (factorial plan 1 x 2) from correspondence analysis of waterbirds abundance (60 species x 4 seasons) in all Jijel eco–complex: A, species factorial map; B, dates. (For the meaning of the species’ codes, see table 1). Fig. 4. Gráfico de ordenación (plan factorial 1 x 2) de un análisis de correspondencia de la abundancia de aves acuáticas (60 especies x 4 temporadas) en el conjunto del eco complejo de Jijel: A, mapa factorial de especies; B, fechas (ver el significado de los códigos de especies, véase la tabla 1).
On the whole, this work reconfirms the Jijel wetlands eco-complex as a particularly important wintering area for Palearctic waterbirds. These environments are used by several hundred waterfowl during seasonal migrations, as well as for wintering, nesting and rearing young. The wetland bird communities in this area are composed of rare and specially protected species such as the ferruginous duck, the white-headed duck, and the shelduck (Tadorna tadorna).
Future research should continue monitoring surveys of rare species, identify trends in their population dynamics, and try to elucidate the relationship between limnological characteristics of wetlands related to anthropogenic influences and pollution. These conditions are likely to be exacerbated by climate change and the assemblage of the waterfowl population. The long–term preservation of this wetlands eco-complex and all other threatened ecosystems in North Africa can only be effective through the establishment of a management plan based on a sustainable compromise between the biodiversity conservation and the needs of the autochthonous populations.
Acknowledgements
The authors wish to thank the many landowners for allowing us to work on their property. We thank the editor and anonymous reviewers for their invaluable comments that helped to substantially improve the manuscript. We also thank Chraitaia H. for the statistical analysis. This research was financially supported by the Algerian Ministry of Higher Education and Scientific Research (Projects PNR and CNEPRU No. F: 01720140025).
















