
Arxius de Miscel·lània Zoològica. Volume 16 (2018) Pages: 173-183
Diversidad de parasitoides primarios y secundarios del pulgón Myzus persicae (Hemiptera, Aphididae) en el duraznero Prunus persica (Rosales, Rosaceae) en la provincia de Mendoza, Argentina
Mazzitelli, M. E., Aquino, D. A., Gallardo, F. E., Reche, V. A., Ricci, E. M.
DOI: https://doi.org/10.32800/amz.2018.16.0173Download
PDFCite
Mazzitelli, M. E., Aquino, D. A., Gallardo, F. E., Reche, V. A., Ricci, E. M., 2018. Diversidad de parasitoides primarios y secundarios del pulgón Myzus persicae (Hemiptera, Aphididae) en el duraznero Prunus persica (Rosales, Rosaceae) en la provincia de Mendoza, Argentina. Arxius de Miscel·lània Zoològica, 16: 173-183, DOI: https://doi.org/10.32800/amz.2018.16.0173-
Reception date:
- 22/05/2018
-
Acceptation date:
- 16/08/2018
-
Publication date:
- 02/10/2018
-
Share
-


-
Visits
- 1151
-
Downloads
- 318
Resumen
Diversidad de parasitoides primarios y secundarios del pulgón Myzus persicae (Hemiptera, Aphididae) en el duraznero Prunus persica (Rosales, Rosaceae) en la provincia de Mendoza, Argentina
Mendoza es la mayor productora de frutas de carozo y la principal elaboradora de durazno para industria de Argentina. Myzus persicae, el pulgón verde o rulo del duraznero, es una de las principales plagas que afecta a este cultivo. Los himenópteros parasitoides de la familia Braconidae destacan como uno de los enemigos naturales más eficientes de esta plaga. Los objetivos del presente trabajo fueron determinar el parasitoidismo natural de este áfido en el duraznero y conocer la diversidad de parasitoides asociados a esta plaga. Durante los años 2015/2016 se realizaron colectas semanales de septiembre a diciembre de brotes con áfidos que presentaban o no señales de parasitoidismo. Se calcularon el porcentaje de parasitoidismo y la frecuencia y abundancia relativa de parasitoides. El porcentaje de parasitoidismo promedio por brote fue cercano al 20 %. Sobre la asociación duraznero–M. persicae, se registraron nueve especies de parasitoides primarios y cuatro especies de parasitoides secundarios. Aphidius colemani fue la especie dominante dentro de los parasitoides primarios.
Palabras clave: Áfidos, Hymenoptera, Braconidae, Aphidiinae, Figitidae, Pteromalidae
Abstract
Diversity of primary and secondary parasitoids of the aphid Myzus persicae (Hemiptera, Aphididae) in peach Punus persica (Rosales, Rosaceae) in the province of Mendoza, Argentina
Mendoza is the highest stone fruit producer and the main industrial peach producer for Argentina. Myzus persicae, known as the green peach aphid or the peach–potato aphid, is one of the main pests that affect this crop. Hymenoptera parasitoids of the Braconidae family stand out as one of the most efficient groups of natural enemies of this pest. The objectives of this study were to determine the natural parasitoidism of this peach aphid and to know the diversity of parasitoids associated with this pest. During the years 2015/2016, weekly collections were made from September to December of shoots with aphids that might or might not show signs of parasitoidism. The percentage of parasitoidism and the frequency and relative abundance of the parasitoids were calculated. The percentage of average parasitoidism per shoot was close to 20 %. About the peach–M. persicae association nine species of primary parasitoids and four species of secondary parasitoids were recorded. Aphidius colemani was the dominant species within the primary parasitoids.
Key words: Aphids, Hymenoptera, Braconidae, Aphidiinae, Figitidae, Pteromalidae
Resum
Diversitat de parasitoides primaris i secundaris del pugó Myzus persicae (Hemiptera, Aphididae) en la bresquillera Prunus persica (Rosales, Rosaceae) a la província de Mendoza, Argentina
Mendoza és la primera productora de fruita de pinyol i la principal elaboradora de bresquilla per a indústria de l’Argentina. Myzus persicae, el “pugó verd”, és una de les principals plagues que afecta aquest conreu. Els himenòpters parasitoides de la família Braconidae destaquen com un dels enemics naturals més eficients d’aquesta plaga. Els objectius d’aquest treball van ser determinar el parasitoidisme natural d’aquest afídid a la bresquillera i conèixer la diversitat de parasitoides associats a aquesta plaga. Durant els anys 2015/2016 es van portar a terme recol·leccions setmanals de setembre a desembre de brots amb afídids que presentaven o no senyals de parasitoidisme. Es van calcular el percentatge de parasitoidisme i la freqüència i abundància relativa de parasitoides. El percentatge de parasitoidisme mitjà per brot va ser de prop del 20 %. Pel que fa a l’associació bresquillera–M. persicae, es van registrar nou espècies de parasitoides primaris i quatre espècies de parasitoides secundaris. Aphidius colemani va ser l’espècie dominant entre els parasitoides primaris.
Paraules clau: Afídids, Hymenoptera, Braconidae, Aphidiinae, Figitidae, Pteromalidae
Introducción
La provincia de Mendoza es la mayor productora de frutas de hueso y la principal elaboradora de durazno Prunus persica (L.) Batsch para la industria de Argentina, ocupando el sexto lugar a nivel mundial (IDR, 2011). Los rendimientos de este cultivo pueden verse afectados, entre otros factores, por problemas sanitarios entre los que se destaca la presencia del pulgón verde o rulo del duraznero Myzus (Nectarosiphon) persicae Sulzer (Hemiptera, Aphididae), debido a los daños económicos que ocasiona (Bentley et al., 1999; Becerra et al., 2011).
En Mendoza, el pulgón verde presenta un ciclo de vida holocíclico dioico realizando su fase sexual en el duraznero, su hospedero primario, mientras que entre los hospederos secundarios se conocen numerosas hortalizas y flora silvestre (Ortego and Carrillo, 1995; Blackman and Eastop, 2000). Entre los daños directos que produce el áfido en el frutal se encuentra el enrulado y distorsión de las hojas apicales (rulo del duraznero) y el retardo del crecimiento de los brotes nuevos; y en el durazno pelón (Prunus persica var. nucipersica) puede atacar los frutos provocando deformaciones y áreas descoloridas (Barbagallo et al., 1998; Cucchi and Becerra, 2006). Por otro lado, entre los daños indirectos se encuentran la producción de fumagina y la transmisión de virus (Barbagallo et al., 1998; Becerra et al., 2011). El virus Plum pox (PPV), causante de la enfermedad de la sharka, es transmitido por distintas especies de pulgones en forma no persistente, destacándose M. persicae por su eficiencia como vector del mismo (Gildow et al., 2004). Ortego et al. (2006) reportaron la presencia de la enfermedad en la zona de Cuyo en el año 2005, quedando circunscrita en la actualidad a diferentes focos en las provincias de San Juan y en el sur de Mendoza (Pizzuolo et al., 2011).
Debido a la magnitud de los daños que ocasiona en la economía regional, es de fundamental importancia el reconocimiento de los enemigos naturales de la plaga presentes en dicho agroecosistema, herramienta fundamental para desarrollar y posteriormente implementar programas de control biológico (Van Driesche et al., 2007). Un grupo importante de enemigos naturales de M. persicae son los parasitoides representantes de la familia Braconidae (Aphidiinae), entre los que destacan las especies Aphidius colemani Viereck, A. ervi Haliday y Diaeretiella rapae (M´Intosh) (Hymenoptera, Braconidae), entre otras (Starý et al., 1993). En Argentina, la información se refiere principalmente A. colemani (De Santis and Esquivel, 1966; De Santis, 1967; Starý and Delfino, 1986; Botto and Hernández, 1989; Ovruski et al., 1998; Berta et al., 2002; Andorno et al., 2007; Loiácono et al., 2008; Zumoffen et al., 2015). En la provincia de Mendoza se registró la presencia de especies de Aphidius Nees, Praon Haliday y Ephedrus Haliday parasitoidizando a M. persicae en el duraznero (Becerra et al., 2011).
Los objetivos del presente estudio fueron: determinar el parasitoidismo natural de
Myzus persicae en el cultivo del duraznero y conocer la diversidad de parasitoides asociados a este áfido en la provincia de Mendoza, Argentina, con miras a su utilización en el control biológico por conservación.
Material y métodos
Los estudios se llevaron a cabo en un monte comercial de producción de duraznero “Bowen” de una hectárea, ubicado en la localidad de La Colonia, departamento de Junín, provincia de Mendoza (33° 06′ 42.9” S, 68° 28′ 40.3” W) durante las temporadas 2015 y 2016. El sistema de conducción empleado en la implantación del monte frutal fue en vaso tradicional y los árboles fueron injertados sobre el portainjerto Garfinem (Prunus persica x Prunus dulcis), a una distancia de 5,5 m entre hileras y 4 m entre plantas, con un total de 450 plantas. Se utilizó como cobertura verde Avena sativa L. y no se realizaron aplicaciones de productos fitosanitarios.
Para la determinación del parasitoidismo natural de M. persicae se realizaron muestreos semanales desde mediados de septiembre hasta principios de diciembre, período de mayor incidencia de la plaga en el cultivo de acuerdo con su ciclo biológico. En cada fecha de muestreo se recolectaron cuatro brotes con presencia de pulgones por planta, en 16 plantas, dando un total de 832 brotes para ambos años de estudio. Las muestras fueron colocadas en bolsas plásticas rotuladas y mantenidas en heladera durante una semana para su posterior observación en el laboratorio. Posteriormente se contabilizó la cantidad de ejemplares que presentaron o no señales de parasitoidismo, es decir pulgones parasitoidizados (momias) y sin parasitoidizar. Las observaciones se realizaron bajo microscopio binocular estereoscópico marca Zeiss modelo Stemi 2000 C.
Con los datos obtenidos se calculó el porcentaje de parasitoidismo como el cociente entre el número de momias y el total de áfidos (pulgones sin parasitoidizar más momias) (Mazzuferi et al., 2011). Las momias fueron separadas de las hojas mediante un pincel, dispuestas individualmente en tubos Eppendorf con tapa de algodón y colocadas en incubadora a 20 °C hasta la emergencia de los parasitoides adultos para su identificación.
Se calculó la frecuencia relativa como la relación porcentual entre la frecuencia de la especie y la suma de frecuencias de todas las especies y la abundancia relativa como la relación porcentual del número de individuos de la especie con respecto al total de individuos de todas las especies (Moreno, 2001). Para la identificación de los parasitoides se emplearon las claves de Botto and Hernández (1989), Starý et al. (1991), Zumoffen et al. (2015), Ferrer–Suay et al. (2012), Gibson et al. (1997), Gibson and Vikberg (1998) y Gibson (2001).
Los ejemplares obtenidos fueron depositados en la colección del Museo de La Plata, Buenos Aires, Argentina, y en la colección entomológica de la Estación Experimental Agropecuaria Junín, Mendoza, Argentina.
Resultados
Myzus persicae fue la única especie de áfido identificada en este estudio. El pico de abundancia de este pulgón se registró, para ambos años, a fines de octubre, coincidente con el estado fenológico de máximo crecimiento de brotes. Los niveles de abundancia de áfidos y parasitoidismo por brote variaron significativamente de un año al otro, siendo notablemente mayores en el segundo año (fig. 1).

Fig. 1. Parasitoidismo natural de Myzus persicae y número de áfidos por brote sobre el duraznero. Temporadas 2015 y 2016. Fig. 1. Natural parasitoidism of Myzus persicae and number of aphyds per shoot on peach trees in 2015 and 2016.
El porcentaje promedio de parasitoidismo en ambos años fue de 19,67 %. En general, cuando el porcentaje de parasitoidismo fue alto, la abundancia de pulgones fue baja y viceversa. Los mayores porcentajes de parasitoidismo se obtuvieron desde mediados de noviembre a principios de diciembre en ambas temporadas de estudio, coincidentemente con bajos niveles de abundancia de M. persicae (fig. 1).
Se colectaron 2.152 momias de las cuales se obtuvieron 1.794 individuos (parasitoides primarios y secundarios). Considerando la emergencia total de parasitoides adultos de las momias en el año 2015, el 29,61 % correspondió a parasitoides primarios y el 70,39 % a parasitoides secundarios (hiperparasitoides). Con respecto al año 2016, el 38,35 % correspondió a parasitoides primarios y el 61,65 % a parasitoides secundarios.
Se identificaron nueve especies de parasitoides primarios asociados a M. persicae, siendo A. colemani la más frecuente, seguida por A. matricariae Haliday (fig. 2). Con respecto a la abundancia, A. colemani fue la especie más abundante, en segundo lugar se encontraron A. avenae Haliday y Praon volucre Haliday. Además, el género Aphidius superó el 70 % de la abundancia total (fig. 2).
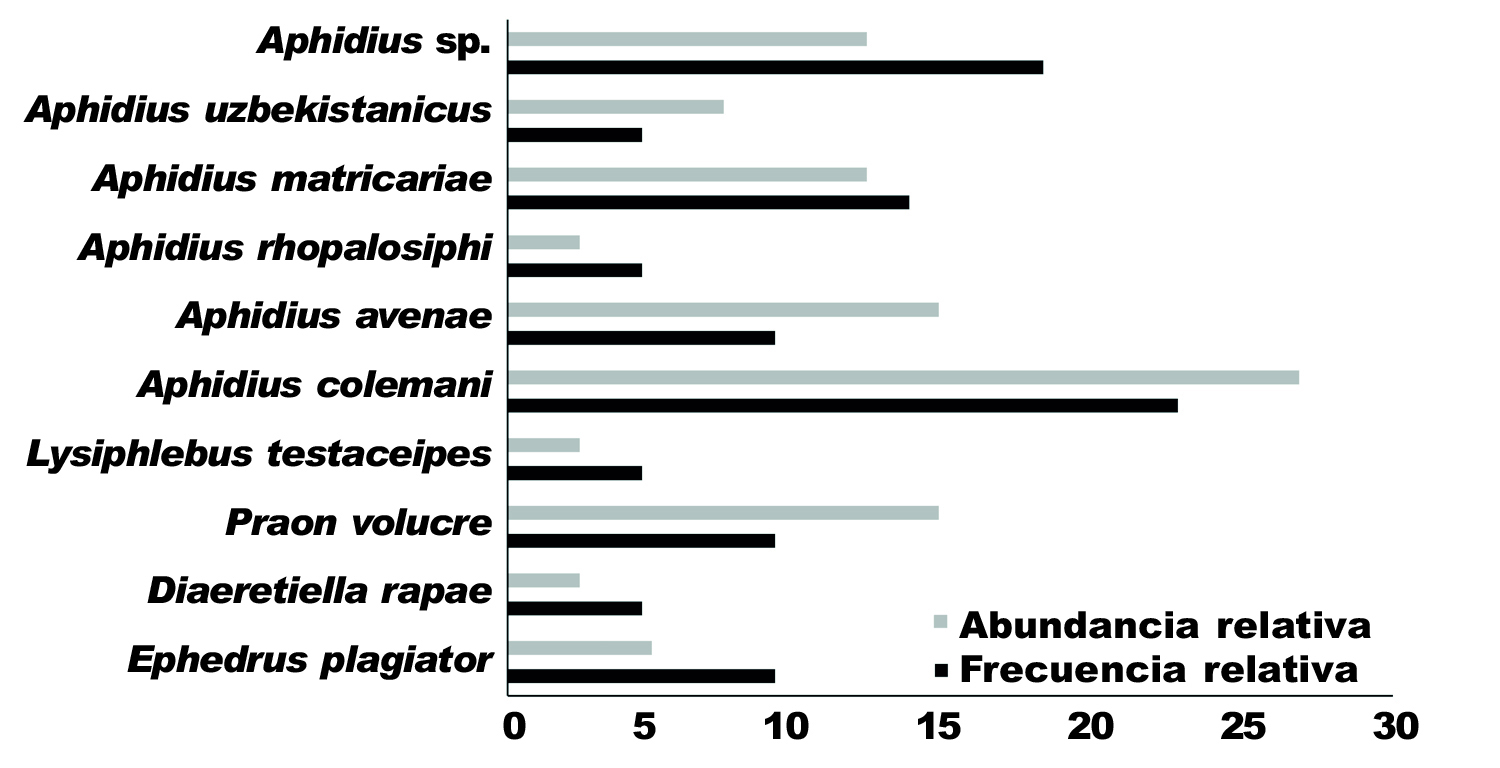
Fig. 2. Porcentaje de frecuencia y abundancia relativa de parasitoides primarios sobre Myzus persicae. Temporadas 2015 y 2016. Fig. 2. Percentage of frequency and relative abundance of primary parasites on Myzus persicae in 2015 and 2016.
Asimismo, podemos observar que la mayor emergencia de los parasitoides primarios se concentró durante los meses de octubre y noviembre. Solamente durante el mes de noviembre se encontró la totalidad de las especies registradas. Aphidius colemani fue la única especie que estuvo presente durante los cuatro meses de muestreo (fig. 3).

Fig. 3. Temporalidad en la emergencia de parasitoides primarios de Myzus persicae. Temporadas 2015 y 2016: Ep, Ephedrus plagiator; Pv, Praon volucre; Lt, Lysiphlebus testaceipes; Ac, Aphidius colemani; Ap, Aphidius picipes; Ar, Aphidius rhopalosiphi; Au, Aphidius uzbekistanicus; Am, Aphidius matricariae; Dr, Diaeretiella rapae. Fig. 3. Months of emergence of Myzus persicae parasites in 2015 and 2016. (For abbreviations of species, see above.)
Con relación a los hiperparasitoides, se encontraron cinco especies, entre las cuales la familia Figitidae (Hymenoptera, Cynipoidea) concentró la mayor parte de los parasitoides obligados asociados al parasitoidismo natural de Myzus persicae, con las especies Alloxysta desantisi (Pujade–Villar and Díaz) y Alloxysta consobrina (Zetterstedt), siendo la primera la más frecuente y abundante. Le siguió en frecuencia y abundancia la especie Pachyneuron aphidis (Bouché), hiperparasitoide facultativo de la familia Pteromalidae (Hymenoptera, Chalcidoidea). También se registraron ejemplares de Asaphes suspensus (Nees), hiperparasitoide obligado (Pteromalidade, Hymenoptera, Chalcidoidea) y representantes del género Dendrocerus Ratzeburg (Hymenoptera, Ceraphronoidea) (fig. 4).

Fig. 4. Porcentaje de frecuencia y abundancia relativa de parasitoides secundarios asociados a Myzus persicae. Temporadas 2015 y 2016. Fig. 4. Percentual frequency and relative abundance of secondary parasites associated with Myzus persicae in 2015 and 2016.
Por otro lado, la mayor parte de los parasitoides secundarios emergieron durante los meses de noviembre y diciembre (fig. 5). La emergencia de los parasitoides secundarios se registró, en ambos años, a partir de los últimos días de octubre. Debido a la metodología utilizada no se pudo identificar a sus huéspedes primarios.

Fig. 5. Temporalidad en la emergencia de parasitoides secundarios asociados a Myzus persicae. Temporadas 2015 y 2016: Pa, Pachyneuron aphidis; As, Asaphes suspensus; D sp., Dendrocerus sp.; Ad, Alloxysta desantisi; Ac, Alloxysta consobrina. Fig. 5. Months of emergence of parasites secondary to Myzus persicae in 2015 and 2016. (For the abbreviations of species, see above.)
Por consiguiente, se dan a conocer por primera vez en la provincia de Mendoza las siguientes cuatro relaciones tróficas: Prunus persica–M. persicae–A. desantisi/A. consobrina/P. aphidis/A. suspensus.
Discusión
El presente trabajo permitió conocer la dinámica en las poblaciones de la plaga Myzus persicae y sus correspondientes parasitoides en el cultivo del duraznero en Junín, Mendoza, Argentina. En este estudio, la tasa de parasitoidismo promedio entre ambos años varió en un rango de entre 4,37–34,98 %, dando un promedio de 19,67 %. Estudios realizados a laboratorio por Ferreira et al. (2018) determinaron una tasa de parasitoidismo sobre M. persicae con variaciones de 4,5–62,2 %. Por su parte, Sampaio et al. (2001) determinaron un parasitoidismo con variaciones de 38–62 % en densidades de M. persicae de 80–20 áfidos. Berta et al. (2002) estimó un porcentaje promedio de parasitodismo de 10,5 % en estudios realizados en tomate a campo, donde M. persicae fue el principal áfido asociado al cultivo.
Con relación a la diversidad de parasitoides primarios, en este estudio se pudo constatar que Aphidius colemani fue la especie más frecuente y abundante. El complejo Aphidius spp. sobresalió en cuanto a la abundancia total de los parasitoides primarios. Por su parte, Berta et al. (2002) destacan en su trabajo la frecuencia y abundancia de las especies Myzus persicae y A. colemani en el cultivo de tomate. Esto también fue corroborado por Andorno (2012), quien destaca a. colemani como el principal parasitoide de M. persicae en el cultivo de rúcula.
Se puede observar que en ambos años, los mayores porcentajes de parasitoidismo se asociaron a bajas poblaciones de M. persicae. Esto es coincidente con la respuesta funcional de A. colemani, al que se le asigna una respuesta funcional de tipo III. En este caso, el número de áfidos parasitoidizados se acelera con el aumento de la densidad de la plaga, hasta un cierto tiempo en que comienza a limitarse (Fernández–Arhex and Corley, 2003). A su vez, valores elevados al final del ciclo de la plaga podrían asociarse también a la presencia de pulgones momificados en fechas anteriores. Por otra parte, los parasitoides primarios emergieron principalmente durante los meses de octubre y noviembre, mientras que los secundarios lo hicieron durante noviembre y diciembre, por lo que los valores tan altos de parasitoidismo encontrados al final de la temporada se debieron probablemente a la presencia de hiperparasitoides, afectando a los valores promedios de parasitoidismo natural. Berta et al. (2002) encontró valores menores de parasitoidismo a campo (10,5 % frente a 19,6 7 % en nuestro estudio). Sin embargo, a diferencia de los resultados obtenidos en ese estudio, el hiperparasitoidismo de M. persicae sobre el duraznero en ambas temporadas desempeñó un importante papel dentro del parasitoidismo natural, ya que del total de ejemplares emergidos desde las momias en ambos años de estudio, alrededor del 60 % en promedio en los años 2015–2016 fueron especies asociadas a parasitoides secundarios. A partir de esto se deduce que el mayor valor promedio de parasitoidismo en nuestro estudio se asocia a una mayor incidencia de parasitoides secundarios. Dichos autores registraron también, como en el trabajo que aquí se presenta, una emergencia posterior de los parasitoides secundarios con relación a los primarios.
Con respecto al hiperparasitoidismo, estudios de Walgenbach (1994), Acebes and Messing (2013), Prado et al. (2015) y Sampaio et al. (2017) determinaron que los hiperparasitoides limitaron la eficiencia de los parasitoides primarios (entre los que se cuenta la especie A. colemani) en el control biológico de M. persicae y otros áfidos por su elevado porcentaje. En el presente trabajo pudimos observar que el desempeño de los parasitoides primarios probablemente se vio afectado principalmente en el mes de noviembre, época de emergencia común de parasitoides primarios y secundarios. Con relación a las especies de parasitoides secundarios asociados a Myzus persicae, se encontraron cuatro citas nuevas en la provincia de Mendoza: Asaphes suspensus, Pachyneuron aphidis, Alloxysta consobrina y Alloxysta desantisi. Estudios similares realizados por Cano et al. (1998), Berta et al. (2002) y Bañol et al. (2012), encontraron a las especies P. aphidis, A. suspensus, Dendrocerus carpenteri (Hymenoptera, Megaspilidae) y Alloxysta sp. como hiperparasitoides de áfidos plaga. En nuestro estudio no se pudieron establecer las especies de parasitoides primarios afectados, aunque probablemente hayan sido las distintas especies del género Aphidius.
Al observar la temporalidad en el emergencia de los parasitoides, podemos constatar que A. colemani destaca por ser el único parasitoide que estuvo presente durante todo el ciclo de la plaga, resaltando su importancia como controlador biológico de M. persicae. Por otra parte, los hiperparasitoides más abundantes, pertenecientes a la familia Figitidae, se concentraron al final del ciclo de la plaga. Esto también fue referido por Berta et al. (2002).
Los resultados que aquí se presentan constituyen una aportación local para el conocimiento de las relaciones tróficas entre Myzus persicae y sus respectivos parasitoides en términos de biodiversidad y para el diseño de estrategias de manejo en el control de esta plaga. La asociación M. persicae–Aphidius colemani resulta de interés para la implementación de prácticas de control biológico en el cultivo de duraznero. Por otra parte, la presencia de parasitoides secundarios encontrados espontáneamente en el agroecosistema es un aspecto a considerar ante la implementación de un plan de control biológico en el cultivo, dado el alto porcentaje de hiperparasitoides identificados. El diseño de una estrategia que asegure una alta presencia de parasitoides primarios al comienzo del ciclo biológico de la plaga es fundamental para lograr el éxito en el control biológico de la misma.
















