
Arxius de Miscel·lània Zoològica. Volum 11 (2013) Pàgines: 181-195
Abundancia y distribución de la liebre ibérica (Lepus granatensis Rosenhauer, 1856) en el Parque Natural de la Sierra de Mariola (Alicante-Valencia)
Belda, A., Arques, J., Peiró, V., Martínez-Pérez, J. E., Zaragozí, B.
DOI: https://doi.org/10.32800/amz.2013.11.0181Descarregar
PDFCita
Belda, A., Arques, J., Peiró, V., Martínez-Pérez, J. E., Zaragozí, B., 2013. Abundancia y distribución de la liebre ibérica (Lepus granatensis Rosenhauer, 1856) en el Parque Natural de la Sierra de Mariola (Alicante-Valencia). Arxius de Miscel·lània Zoològica, 11: 181-195, DOI: https://doi.org/10.32800/amz.2013.11.0181-
Data de recepció:
- 18/11/2013
-
Data d'acceptació:
- 18/03/2014
-
Data de publicació:
- 21/04/2014
-
Compartir
-


-
Visites
- 4396
-
Descàrregues
- 1148
Resumen
Abundancia y distribución de la liebre ibérica Lepus granatensis Rosenhauer, 1856) en el Parque Natural de la Sierra de Mariola (Alicante-Valencia)
La liebre ibérica (Lepus granatensis Rosenhauer, 1856) es una especie de gran valor en los ecosistemas mediterráneos españoles por varios motivos: por su interés cinegético, por servir de alimento a numerosos predadores, por mejorar la fertilidad del suelo e incrementar la diversidad vegetal, etc. Sin embargo, factores como la fragmentación, la degradación y la pérdida de hábitats, así como las enfermedades, la predación y la excesiva presión cinegética que afectan a la especie, están influyendo de manera negativa en su conservación. Por esta razón resulta interesante conocer la abundancia y distribución de la liebre ibérica en diferentes zonas de la península de las que no se tienen suficientes datos, para que el análisis de su conjunto proporcione pautas que permitan realizar una gestión adecuada para mantener e incrementar las poblaciones de liebres en la península. Nuestro objetivo es conocer la abundancia y distribución de la liebre ibérica en los principales usos del paisaje de una zona de montaña del Mediterráneo peninsular, el Parque Natural de la Sierra de Mariola, situado entre las provincias de Alicante y Valencia, durante diferentes periodos de su ciclo biológico anual. Los datos obtenidos en los transectos realizados entre 2008 y 2010 muestran que las mayores abundancias anuales tienen lugar en primavera (IKA medio de 0,26 liebres/km) y disminuyen en invierno (IKA medio de 0,075 liebres/km). En cuanto a sus preferencias relacionadas con los diferentes usos del suelo, las mayores abundancias se obtienen en la matriz de cultivos de secano (IKA medio de 0,33 liebres/km) y en la de cultivos de regadío (IKA medio de 0,2 liebres/km). Las matrices de vegetación natural y abandono agrícola presentan bajo número de liebres, con valores de 0,083 y 0,033 liebres/km respectivamente.
Palabras clave: Abundancia, IKA (índice kilométrico de abundancia), Lepus granatensis, Matriz del paisaje, Sierra de Mariola
Abstract
Abundance and distribution of the Iberian hare (Lepus granatensis Rosenhauer, 1856) in the Sierra de Mariola Natural Park (Alicante-Valencia)
The Iberian hare (Lepus granatensis Rosenhauer, 1856) is a species of great value in Spanish Mediterranean ecosystems for several reasons, such as its interest to hunters, its contribution to soil fertility and plant diversity, and its role as prey. However, factors such as fragmentation, degradation and loss of habitat and diseases, predation and high pressure hunting are having a detrimental effect on the conservation of the species. It is therefore of interest to determine the abundance and distribution of the Iberian hare in areas of the peninsula where there are insufficient data to establish guidelines for conservation and sustainable management of hare populations in the peninsula. Our goal was to assess the abundance and distribution of the Iberian hare in the most widely used areas of a mountainous Mediterranean landscape in the Iberian peninsular (Mariola Mountain Park, located between the provinces of Alicante and Valencia). Data obtained from studies conducted in transects from 2008 to 2010 showed that intra-annual abundance was highest in spring (KIA half of 0.26 hares/km) and lowest in winter (average 0.075 KIA hares/km). As to their preferences in relation to land use, abundance was highest in the matrix of dry groves (KIA half of 0.33 hares/km) and irrigated groves (average 0.2 KIA hares/km). The matrix of natural vegetation and agricultural abandonment had low numbers of hares, with values of 0.083 and 0.033 hares/km, respectively.
Key words: Abundance, KAI (Kilometric Abundance Index), Lepus granatensis, Landscape matrix, Sierra de Mariola
Resum
Abundància i distribució de la llebre ibèrica (Lepus granatensis Rosenhauer, 1856) al Parc Natural de la Serra de Mariola (Alacant-València)
La llebre ibèrica (Lepus granatensis Rosenhauer, 1856) és una espècie de gran valor als ecosistemes mediterranis espanyols per diversos motius: per l’interès cinegètic, perquè serveix d’aliment a nombrosos predadors, perquè millora la fertilitat del sòl i incrementa la diversitat vegetal, etc. Tanmateix, factors com ara la fragmentació, la degradació i la pèrdua d’hàbitats, com també les malalties, la predació i la pressió cinegètica excessiva que afecten l’espècie estan influint de manera negativa en la seva conservació. És per això que resulta interessant conèixer l’abundància i distribució de la llebre ibèrica en diferents zones de la Península de les quals no es tenen prou dades a fi que l’anàlisi del conjunt proporcioni pautes que permetin una gestió adequada per mantenir i incrementar les poblacions de llebres a la Península. El nostre objectiu és conèixer l’abundància i distribució de la llebre ibèrica en els principals usos del paisatge d’una zona de muntanya del Mediterrani peninsular, el Parc Natural de la Serra de Mariola, situat entre les províncies d’Alacant i València, durant diferents períodes del seu cicle biològic anual. Les dades obtingudes als transsectes fets entre 2008 i 2010 mostren que les abundàncies anuals més grans tenen lloc a la primavera (IKA mitjà de 0,26 llebres/km) i disminueixen a l’hivern (IKA mitjà de 0,075 llebres/km). Pel que fa a les preferències relacionades amb els diferents usos del sòl, les abundàncies més grans s’obtenen a la matriu de conreus de secà (IKA mitjà de 0,33 llebres/km) i a la de cultius de regadiu (IKA mitjà de 0,2 llebres/km). Les matrius de vegetació natural i abandonament agrícola presentes un nombre de llebres baix, amb valors de 0,083 i 0,033 llebres/km, respectivament.
Paraules clau: Abundancia, IKA (índex quilomètric d’abundància), Lepus granatensis, Matriu del paisatge, Sierra de Mariola
Introducción
La liebre ibérica (Lepus granatensis Rosenhauer, 1985) es una especie de gran importancia en los ecosistemas de la península Ibérica porque sirve de alimento a un buen número de depredadores (Carro, 2005; Carro & Soriguer, 2010) hecho que se ha acentuado en los últimos años debido sobre todo al considerable descenso poblacional del conejo de monte (Oryctolagus cuniculus). Además, contribuye a mejorar la fertilidad del suelo y a incrementar la diversidad vegetal, igual que el conejo de monte, al dispersar las semillas de los frutos que consume (Galvez et al., 2008). También, es muy importante, desde el punto de vista económico, ya que es una pieza muy codiciada por los cazadores (Duarte & Vargas, 1998; Vargas, 2002).
La liebre ibérica presenta ciertas adaptaciones fisiológicas al medio (Corbet, 1983) y una elevada capacidad reproductiva (Alves et al., 2002; Fernández et al., 2008) que le han permitido ocupar buena parte de la península y adaptarse a una gran variedad de hábitats, desde zonas secas como dunas costeras, hasta zonas de montaña húmeda (Palacios, 1978; Palacios & Meijide, 1979; Carro & Soriguer, 2002). Las liebres tienen la capacidad de adaptar su dieta a las condiciones del hábitat en el que se encuentran y a las circunstancias de cada momento (Chapuis, 1990; Carro, 2005). La selección de hábitat varía con la época del año ya que la especie utiliza cada medio en el momento fenológico en el que este satisface sus necesidades, en especial las alimentarias (Pepin, 1985; Tapper & Barnes, 1986). Los requerimientos alimentarios pueden cubrirse en medios diversificados (Duarte, 2000). Las liebres son animales de actividad crepuscular y nocturna con capacidad para recorrer grandes distancias (Wray, 1992) y, por lo general, conocen minuciosamente el territorio que habitan, lo que les permite, en numerosas ocasiones, huir de los predadores.
Las poblaciones de liebre ibérica presentan una distribución desigual de la abundancia en la península Ibérica, de manera que en amplias zonas del norte de peninsular ha desaparecido o se encuentra en declive (Palacios & Ramos, 1979; Ballesteros et al., 1996; Duarte, 2000), mientras que en otras zonas, como Andalucía,su situación es estable, con tendencia incluso a la expansión desde la década de los años 1980 (Duarte & Vargas, 1998; Carro, 2005).
El objetivo general de este trabajo es conocer la abundancia y distribución de la liebre ibérica en una zona de media montaña del mediterráneo peninsular, el Parque Natural de la Sierra de Mariola. El estudio se ha realizado entre octubre de 2008 y mayo de 2010 con el propósito de abarcar los siguientes objetivos específicos: (1) analizar la variación temporal de la abundancia de la especie, según los periodos de su ciclo biológico anual, relacionados principalmente con los periodos reproductor y cinegético, (2) analizar la variación temporal de la abundancia de la especie a lo largo de los tres años de estudio y comparar los datos con otras zonas de la península y (3) analizar el grado de influencia de las principales matrices del paisaje en la abundancia de la especie.
En general, este trabajo permitirá tener un conocimiento más preciso de los factores ecológicos (variables del hábitat) que intervienen en la distribución y abundancia de las liebres, información que puede ser utilizada para mejorar el manejo y conservación de la especie en zonas montañosas del mediterráneo peninsular y, en concreto, pueden servir como recomendaciones prácticas para realizar una gestión adecuada dirigida a mantener y aumentar el tamaño de sus poblaciones.
Material y métodos
Área de estudio
El estudio se ha desarrollado en el Parque Natural de la Sierra de Mariola, de 17.500 ha y situado en la Comunidad Valenciana, entre el norte de la provincia de Alicante y el sur de la provincia de Valencia, abarcando parte de las comarcas del Alcoià, la Vall d’Albaida y el Comtat (fig. 1).

Fig. 1. Localización del Parque Natural de la Sierra de Mariola.
Fig. 1. Location of the Sierra de Mariola Natural Park.
El marco físico define un espacio con predominio de zonas montañosas elevadas, de relieves escarpados, en el que se mezclan retablos elevados de materiales permeables (calizos) y valles impermeables (margas miocenas), con alternancia sinclinal-anticlinal característica del estilo jurásico (Belda & Bellod, 2007). Su clima es típicamente mediterráneo con temperaturas suaves, lluvias concentradas en primavera y otoño y un destacado periodo seco en verano. La vegetación climácica del termotipo mesomediterráneo y ombrotipo subhúmedo es el carrascal (asociación Hedero helicis-Quercetum rotundifoliae subas. ulicerosum parviflorae). En cuanto a los usos del suelo actuales (fig. 2), el más abundante configura una matriz natural que ocupa el 67% del total de la superficie del parque, seguido de las áreas ocupadas por el secano (24%) y otros usos como el urbano (5%), los cultivos abandonados (3%) y el regadío (1%) (fig. 3). Hay que señalar que en algunas zonas del Parque Natural se han producido algunos episodios de cambios de uso del suelo destacables, especialmente por la transformación de uso natural en suelo urbano.
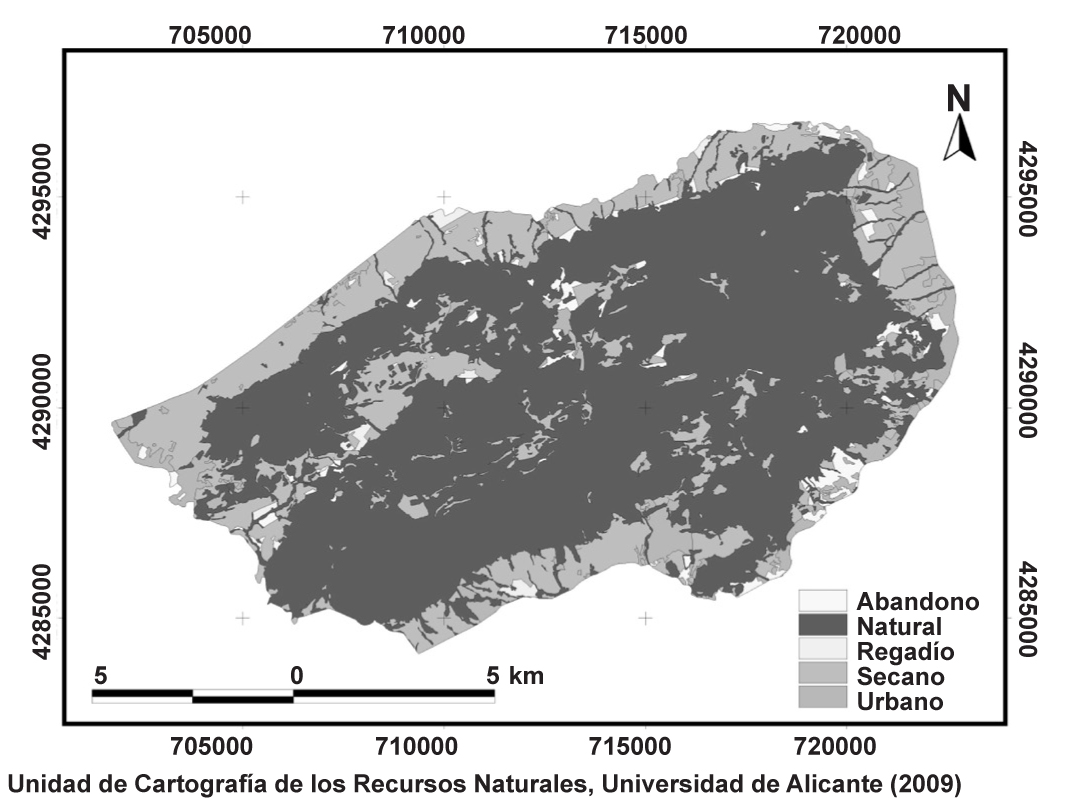
Fig. 2. Distribución de los usos del suelo en el Parque Natural de la Sierra de Mariola.
Fig. 2. Patterns of land use in the Sierra de Mariola Natural Park.

Fig. 3. Porcentaje de superficie de los usos del suelo en el Parque Natural de la Sierra de Mariola.
Fig. 3. Surface percentages of land use in the Sierra de Mariola Natural Park.
Los procesos de urbanización son reconocidos globalmente como una amenaza significativa para la biodiversidad. No obstante, este espacio natural posee un alto valor de diversidad animal y vegetal. Las especies cinegéticas más representativas del parque son el conejo, la perdiz (Alectoris rufa), la paloma torcaz (Columba palumbus), la liebre (Lepus granatensis), el muflón (Ovis musimon), el arruí (Ammotragus lervia), el gamo (Dama dama), el jabalí (Sus scrofa) y el zorro (Vulpes vulpes) (Belda et al., 2009; Arques et al., 2010, 2011).
Digitalización de usos del suelo
Los mapas de usos del suelo se han obtenido de la digitalización y georreferenciación de los fotogramas aéreos correspondientes al año 2005 (escala 1:5.000) del Instituto Cartográfico Valenciano (ICV), mediante la utilización de los programas ER-Mapper® y CartaLinx®. Las coberturas obtenidas han sido exportadas a los formatos de ArcGIS® e Idrisi Andes® para la construcción de la base cartográfica y la obtención de mapas temáticos con el SIG. Asimismo, se han obtenido las coberturas de los núcleos urbanos y las vías de comunicación por cada uno de los cotos, utilizando como fuente la base cartográfica de la Consejería de Obras Públicas (COPUT) de la Generalitat Valenciana. De esta forma se tiene información detallada de los usos del suelo, vías de comunicación, altitud, orientación, pendientes, hidrología, litología y núcleos urbanos de la zona de estudio. Esta cartografía ha sido básica para la selección de los puntos de muestreo.
Se han digitalizado 1.213 polígonos que cubren en total una superficie próxima a las 17.000 ha. Los usos del suelo se representan de forma generalizada: abandono, natural, secano, regadío y urbano (tabla 1).

Tabla 1. Superficie de las diferentes matrices de usos del suelo presentes en el área de estudio. Table 1. Size of land use types in the study area.
Determinación de abundancias
Las abundancias relativas de liebre se han obtenido a partir de transectos estratificados de 1.000 m de longitud. En total, se han realizado 240 itinerarios de censo, a razón de 10 censos en cada uno de los distintos usos del suelo y repetido cada temporada desde octubre de 2008 a mayo de 2010. Cada año se han realizado tres campañas de muestreo: poscinegética o prerreproductora (invierno), posreproductora (primavera) y precinegética (otoño). Dichos transectos se han llevado a cabo, de acuerdo con la tipología del mosaico del paisaje propuesta por Jiménez-García (2007), abarcando la diversidad de matrices de usos del suelo (natural, abandono, secano, regadío y urbano). Los datos de los censos de campo se presentan, para cada una de las matrices de usos del suelo y temporada de muestreo, mediante el cálculo del (IKA) índice kilométrico de abundancia (Ferry & Frochot, 1958).
Resultados
La abundancia intraanual de la liebre ibérica en el parque natural de la Sierra de Mariola varía en las distintas épocas del año, aunque sus diferencias no son significativas (F(2,23) = 3,326, P > 0,05). Durante el periodo de muestreo se ha observado que en los meses de febrero (invierno), los censos han dado un IKA medio de 0,075 liebres/km. Su presencia aumenta en los meses de mayo (primavera), con un IKA medio de 0,26 liebres/km, y vuelve a descender en los meses de octubre (otoño), con un valor de 0,15 liebres/km (anexo 1).

Anexo 1. Datos de campo de la liebre ibérica en el Parque Natural de la Sierra de Mariola. Annex 1. Field data of Iberian hare in the Sierra de Mariola Natural Park.
En cuanto a la abundancia de la liebre a lo largo del periodo de estudio, en 2008 se realizaron solamente censos en el mes de octubre y el IKA obtenido fue de 0,2 liebres/km. En 2009 se realizaron censos en los tres periodos (febrero, mayo y octubre) y el IKA medio anual fue de 0,15 liebres/km. Por último, en 2010 se censó en febrero y mayo y el IKA medio anual fue de 0,16 liebres/km. Si consideramos dos periodos, uno que comprende octubre de 2008 y febrero y mayo de 2009 y, el otro octubre de 2009 y febrero y mayo de 2010, los datos indican que no existen diferencias significativas en la abundancias de liebres entre ambos periodos (F(1,23) = 0,684, P > 0,05). El IKA medio obtenido en los transectos realizados en la zona de estudio entre octubre de 2008 y mayo de 2010 ha sido de 0,16 liebres/km (anexo 1).
Los censos realizados en diferentes puntos de la zona de estudio muestran diferencias significativas entre la abundancia de liebres y las diferentes matrices de los usos del suelo presentes (F(3,23) =7,036, P < 0,05). La mayor abundancia se obtiene en la matriz de cultivos de secano (IKA medio de 0,33 liebres/km), seguida de la matriz de cultivos de regadío (IKA medio de 0,2 liebres/km). Las matrices de vegetación natural y abandono presentan el menor número de individuos, con valores medios de IKA de 0,083 y 0,033 liebres/km respectivamente (fig. 4). El test de Bonferroni muestra diferencias significativas entre las zonas de secano y los cultivos abandonados (P = 0,03) y entre los cultivos de secano y la matriz natural (P = 0,015).

Fig. 4. Índice kilométrico de abundancia (IKA) de liebres ibéricas en la Sierra de Mariola.
Fig. 4. Kilometric abundance index (KAI) of Iberian hares in the Sierra de Mariola.
Discusión
Las poblaciones de liebres en la península ibérica no permanecen estáticas a lo largo del tiempo, ya que están ligadas a cambios estacionales que, a su vez, inciden sobre el alimento y el medio afectando a las tasas de mortalidad y natalidad de la especie (Carro & Soriguer, 2010). El análisis de los datos obtenidos en el presente estudio indica que la abundancia de liebres varía a lo largo del año. La mayor presencia se produce en primavera, desciende en otoño y baja mucho más en invierno. La mayor abundancia de liebres en primavera se debe a varias razones. El periodo de caza finaliza en enero y en los meses posteriores se dan las condiciones adecuadas que propician un incremento reproductor de la especie. Es un periodo en el que hay mayor presencia de alimento debido a las lluvias de otoño e invierno y que, unido a un rango de temperaturas y a unas características determinadas de la luz solar, favorece el aumento del porcentaje de hembras gestantes. Rau et al. (1987) afirman que la calidad del hábitat y la disponibilidad de alimento están directamente relacionadas con el éxito reproductor y con la supervivencia de la especie. Alonso et al. (1997) indican que el mayor porcentaje de hembras preñadas se produce en febrero-abril y en junio-julio. Fernández et al. (2008), afirman que en Navarra la máxima reproducción anual tiene lugar en marzo-abril y mayo-junio. Por otro lado, Alves et al. (2002) indican que la máxima reproducción anual de la liebre ibérica se produce en enero-febrero y julio-agosto.
En otoño, la abundancia de liebres desciende. Los máximos anuales del proceso reproductor tienen lugar entre febrero y julio, dependiendo de las zonas y de las condiciones climáticas. En los meses siguientes, el porcentaje de liebres jóvenes que mueren es considerable. Duarte (2000) indica que la mortalidad juvenil es de un 40% y Carro et al. (1999) estiman que la supervivencia de las liebres adultas oscila entre el 20-30%.
Las menores abundancias anuales de liebres en el parque de la Sierra de Mariola, se dan en los meses de invierno. López et al. (1996) también observaron, en una zona de cultivo extensivo de cereal en la provincia de León, una disminución de las abundancias entre octubre y marzo, atribuida a la explotación cinegética del área. Es un periodo inmediatamente posterior a la finalización del periodo de caza, que se extiende desde mediados de octubre a principios de enero.
La abundancia anual de liebres en los tres años que ha durado el estudio ha sido bastante similar, con valores que oscilan entre 0,15 y 0,2 liebres/km. El número de liebres puede variar en función de varios parámetros: aparición de enfermedades, condiciones climáticas adversas, falta de alimento, manejos inadecuados, alteraciones del medio, presencia de ganado, aumento de predadores oportunistas, sobreexplotación cinegética, furtivismo, etc. En la comarca de la Marina Baja (Alicante) los valores medios de las liebres cazadas entre 1988 y 2008 fueron diferentes, oscilando entre 0,07 liebres/superficie de la temporada de 1992-1993 y 0,001 liebres/superficie de las cuatro últimas temporadas analizadas (CMAAUV, 2008). Por otra parte, la abundancia de liebres en Doñana en el periodo comprendido entre 1995-1998 fue diferente, con valores de IKA que oscilaron entre 1,84 liebres/km de 1995, 0,16 de 1996, 0,4 de 1997 y 1,24 liebres/km del año siguiente. El descenso del número de liebres en los años centrales se debió fundamentalmente a las inundaciones que se produjeron en 1996 y 1997 (Carro et al., 2001). Gortázar et al. (2007) observaron también, en diferentes localidades de Aragón, diferencias significativas en las abundancias de liebres entre 1992 y 2002.
La abundancia media de liebres en el periodo de realización del presente estudio es de 0,16 liebres/km. La abundancia de liebres en esta zona de montaña, situada entre las provincias de Alicante y Valencia, es superior a la de las zonas del noroeste peninsular (sur de Orense) e inferior a otras zonas de la península como la comarca de la Marina Alta (Alicante), Doñana, Aragón, Navarra y zonas de montaña del centro de Portugal (Ensemil y Santinha) (ver tabla 2).

Tabla 2. Valores de IKA en distintas zonas de la península Ibérica. Table 2. KAI values in the different regions of the Iberian peninsula.
En cuanto a la abundancia de liebres según los usos del suelo, la mayor se ha obtenido en la matriz de cultivos de secano, seguida de la matriz de regadío. Las matrices de vegetación natural y abandono son las que han presentado un número inferior de individuos. La matriz de secano del parque está formada por cultivos de olivos y almendros, en las zonas más elevadas, y por girasol y trigo en los valles. Las superficies que ocupan los cultivos de secano son reducidas y forman parches próximos a la matriz natural. Suelen ser áreas manejadas por el hombre con presencia permanente de especies herbáceas. Este tipo de hábitat suele ser el mencionado por numerosos autores como el preferido por la especie. Así, las liebres muestran preferencia por lugares llanos y secos, como cultivos de cereal y olivares de campiña (Moreno et al., 1998). Las tierras aradas, con renovación permanente de la vegetación herbácea y parches de arboleda, son los medios favorables para las liebres (Meriggi & Alieri, 1989, Mariggi & Verri, 1990; Stoate & Tapper, 1993; Mclaren et al., 1997). En Andalucía, la liebre, es especialmente abundante en los cultivos de olivar, girasol y viñedo (Duarte & Vargas, 1998). Autores como Pepin (1985) y Tapper & Barnes (1986) indican que la selección de hábitat varía con la época del año, ya que la especie utiliza cada medio en el momento fenológico en el que este satisface sus necesidades, en especial las alimentarias. De este modo, las liebres se alimentan de herbáceas, sobre todo gramíneas y leguminosas. A finales de verano y principios de otoño incorporan a su dieta semillas y algunos frutos. En invierno también se alimentan de cortezas y hojas de matorrales leñosos (Péroux, 1995; Ballesteros, 1998). Por otra parte, la elección de los espacios abiertos, con vegetación de baja altura, desempeña un papel decisivo en el comportamiento antipredatorio de la liebre, ya que esta especie tiene buena vista y escapa a la carrera de los depredadores (Hewson, 1977).
La matriz que ocupa el segundo lugar en abundancia de liebres es la de cultivos de regadío, a pesar de ocupar solamente un 1% de la superficie del parque. Generalmente son cultivos con riego por goteo, como el olivo y la viña, además de huertos de maíz situados en zonas llanas. El uso continuo del sistema de goteo en la zona de cultivos permite la presencia de hierba fresca en el lugar donde cae el agua y, por lo tanto, hay un aporte de alimento con buen contenido hídrico y proteico listo para ser consumido. En general, la abundancia de liebres está relacionada con los cultivos intensivos de regadío y en estas zonas se observa incluso un aumento de sus densidades (López et al., 1996; Palacios & Ramos, 1979). Además, algunos autores han comprobado que existen mayores densidades de liebres en los medios agrícolas intensivos que en los extensivos (Calzada & Martínez, 1994; López et al., 1996). En la comarca de la Marina Alta (noreste de la provincia de Alicante), la mayor presencia de liebres se obtiene en esta matriz (IKA medio de 1,25 liebres/km), prefiriendo zonas llanas con vegetación abierta o baja y cultivos de regadío, principalmente naranjos (Jiménez-García, 2007). Por otra parte, los cultivos de regadío permiten la presencia de hierba fresca a lo largo de todo el año, proporcionando una heterogeneidad espacial importante en los desplazamientos nocturnos (Chapuis, 1990). En Navarra, las zonas que presentan mayores abundancias (5 liebres/km) se encuentran a lo largo del eje del río Ebro, en localidades con cultivos de regadío (Carro & Soriguer, 2010).
La matriz natural es la que ocupa la mayor superficie del parque y, en cambio, la presencia de liebres en la misma es baja. La matriz natural, en la zona de estudio, se caracteriza por ser bastante homogénea, con pinares maduros, encinares y carrascales, todos con abundante matorral. Además, hay algunas áreas sin arbolado y con matorral postincendio debido a los incendios forestales ocurridos durante la década de los años 1990. En esta zona, la abundancia de liebres es menor porque esta especie requiere la mayor cantidad de recursos en áreas relativamente reducidas (Bray et al., 1997). Jiménez-García (2007) también constata que en zonas de matorral con arbolado desaparecen algunas especies cinegéticas, entre ellas la liebre. La matriz natural es homogénea y no permite la presencia de áreas de transición o bordes, estructuras del paisaje fundamentales para detectar a las especies de interés cinegético, ya que se trata de áreas utilizadas para alimentación o reposo (Jiménez-García, 2007).
Por último, la matriz de cultivos abandonados sólo cuenta con una superficie del 3% del parque y es la que presenta la menor abundancia de liebres. Son zonas de frutales (almendros y olivos) que hace años dejaron de cuidarse por su baja rentabilidad económica. En la actualidad, se mantiene la misma estructura que antaño, con los árboles y abundante matorral. Jiménez-García (2007) ha comprobado que en la comarca de la Marina Alta, en las zonas de matorral con arbolado, las liebres están ausentes. Por otra parte, su presencia está asociada a las áreas de cultivo en continuo manejo y con recambio de especies herbáceas (Meriggi & Verry, 1990; McLaren et al., 1997; Smith et al., 2005) y asociada a la actividad humana (Jiménez-García, 2007), condiciones que no suelen darse en las zonas de abandono.
Conclusión
La cuenca mediterránea es una región de Europa que cuenta con una elevada diversidad de especies y un alto grado de endemismos. Sin embargo, la modificación y pérdida de hábitats están influyendo de manera negativa en la conservación de las especies. Así, los cambios en el uso de la agricultura son sugeridos para explicar el declive de numerosas especies (Palma et al., 1999; Chamberlain & Fuller, 2000; Smith & Bruun, 2002). La liebre ibérica es una especie de gran valor en los ecosistemas mediterráneos españoles por lo que su conocimiento ecológico y su relación con los factores que influyen en la heterogeneidad espacial son esenciales para su conservación y manejo.
En el Parque Natural de la Sierra de Mariola, la liebre está presente de forma continuada pero con diferencias notables según la época del año. Los mayores IKA se obtienen en los
meses de mayo, bajan en octubre y sufren un acusado descenso en los meses de febrero. En cambio, apenas hay diferencias en la abundancia de individuos en los tres años que ha durado el presente estudio. La abundancia de liebres en la Sierra de Mariola es baja y tan solo supera la de ciertas zonas del norte y del noroeste peninsular. Los resultados muestran que en la Sierra de Mariola las liebres están presentes en todas las matrices de usos del suelo, siendo más abundantes en los cultivos de secano y regadío, en detrimento de las zonas abandonadas y naturales. Así, se constata que la abundancia y distribución de la liebre en esta zona está relacionada fundamentalmente con paisajes diversificados (heterogeneidad de parches) en los que en poco espacio encuentran respuesta a los requerimientos básicos para su supervivencia.
La selección del hábitat tiene por objeto satisfacer los requerimientos de la fauna concernientes a la alimentación, refugio y reproducción (Bond et al., 2002). La capacidad de desplazamiento de las liebres entre diferentes hábitats es bien conocida (Pepin, 1985; Andre et al., 1997) y tienen preferencia por las zonas de cultivos (Palacios & Meijide, 1979; Lewandowski & Nowakowski, 1993), los hábitats con protección (Flux & Angermann, 1990), los campos de cultivo que alternan con prados y bosques (Homolka, 1986), las zonas de regadío (Jiménez-García, 2007) y los sistemas agrícolas intensivos (Palacios & Ramos, 1979; López et al., 1996). El manejo del hábitat es la principal estrategia para conservar la vegetación natural y aumentar las zonas de transición donde se establecen los ecotonos. La cobertura que proporciona la estructura de ecotono, con cultivos donde se desarrollan manchas de hierba fresca, minimiza el riesgo de depredación y permite a las especies, como las liebres, optimizar su conducta espacial y acceder fácilmente al alimento y a los refugios. Estos son, pues, los lugares ideales para el establecimiento de nuevas poblaciones o aumentar el tamaño de las ya existentes.
Agradecimientos
A la Dirección Territorial de Caza y Pesca de la Consejería de Territorio y Vivienda de la Provincia de Alicante. A la Dirección y personal del Parque Natural de la Sierra de Mariola. Al Servicio de Protección de la Naturaleza de la Guardía Civil (SEPRONA). A todos los informantes y gestores cinegéticos que desinteresadamente han colaborado en la elaboración de este trabajo. A la Fundación C. V. Victoria Laporta Carbonell que gestiona la Finca de Buixcarró, especialmente a Vicente y Carlos. Este proyecto ha sido financiado en parte por la convocatoria pública de proyectos de investigación para grupos precompetitivos de la Consejería de Educación de la Generalitat Valenciana y por las ayudas para la investigación del Instituto Alicantino de Cultura Juan Gil-Albert.
















