
Arxius de Miscel·lània Zoològica. Volum 19 (2021) Pàgines: 183-196
Population dynamics, seasonal fluctuations and spatial distribution of the olive psyllid Euphyllura olivina Costa (Homoptera, Psyllidae) in Algeria
Guessab, A., Elouissi, M., Lazreg, F., Elouissi, A.
DOI: https://doi.org/10.32800/amz.2021.19.0183Cita
Guessab, A., Elouissi, M., Lazreg, F., Elouissi, A., 2021. Population dynamics, seasonal fluctuations and spatial distribution of the olive psyllid Euphyllura olivina Costa (Homoptera, Psyllidae) in Algeria. Arxius de Miscel·lània Zoològica, 19: 183-196, DOI: https://doi.org/10.32800/amz.2021.19.0183-
Data de recepció:
- 05/03/2021
-
Data d'acceptació:
- 24/08/2021
-
Data de publicació:
- 12/10/2021
-
Compartir
-


-
Visites
- 3116
-
Descàrregues
- 0
Abstract
Population dynamics, seasonal fluctuations and spatial distribution of the olive psyllid Euphyllura olivina Costa (Homoptera, Psyllidae) in Algeria
The olive psyllid Euphyllura olivina Costa (Homoptera, Psyllidae) causes considerable damage in olive groves in the Mediterranean basin, there by affecting production. To assess the rate of infestation and provide elements of integrated pest management, we monitored the life cycle and bio–ecology of this pest through population counts on the leaves and twigs of the tree from 29 March 2019 to 27 March 2020 at two relatively distant olive farms (Ain fares and Oued taria) in the wilaya of Mascara, Algeria. Our results showed that infestation was high on trees in the Oued taria farm. We found that E. olivina developed two generations per year, in spring, influenced by the climatic factors. The difference between the relative abundance of the different life stages (eggs, larval instar 1, 2, 3, 4 and 5, female and male) in the cardinals orientations was significant. Infestation was highest on the southern and central orientations of the tree. Populations fluctuations were significantly affected by season at both farms.
Key words: Bio–ecology, Euphyllura olivina, Phenology, Population dynamics, Olive tree groves, Olive tree pests
Resumen
Dinámica de la población, fluctuaciones estacionales y distribución espacial del psílido del olivo Euphyllura olivina Costa (Homoptera, Psyllidae) en Argelia
El psílido del olivo, Euphyllura olivina Costa (Homoptera, Psyllidae), causa daños considerables en los olivares de la cuenca mediterránea, por lo que afecta a la producción. Para evaluar la tasa de infestación y proporcionar elementos de manejo integrado de plagas, el monitoreo del ciclo de vida y la bioecología de estas plagas se llevaron a cabo a través de conteos poblacionales periódicos en las hojas y ramitas del árbol del 29 de marzo de 2019 al 27 de marzo de 2020 en dos fincas de olivos relativamente distantes (Ain fares y Oued taria) en la wilaya de Mascara, Argelia. Nuestros resultados mostraron que la infestación es alta en los árboles de la finca de Oued taria. El estudio de la bioecología de E. olivina demostró que esta plaga desarrollaba dos generaciones al año, en primavera, en función de los factores climáticos. Se encontró una diferencia significativa entre la abundancia relativa de las diferentes etapas vitales (huevos, fase larvaria 1, 2, 3, 4 y 5, hembras y machos) y las orientaciones de los puntos cardenales. Las orientaciones sur y centro del árbol fueron las más infectadas por psyla. Las fluctuaciones de las poblaciones se vieron significativamente afectadas por las estaciones en las dos fincas.
Palabras clave: Bioecología, Euphyllura olivina, Fenología, Dinámica poblacional, Olivares, Plagas en olivares
Resum
Dinàmica de la població, fluctuacions estacionals i distribució espacial del psíl·lid olivar Euphyllura olivina Costa (Homoptera, Psyllidae) a Argèlia
El psíl·lid de l’olivera, Euphyllura olivina Costa (Homoptera; Psyllidae), provoca danys considerables als camps d’oliveres de la conca mediterrània, per tant n’afecta la producció. Per avaluar la taxa d’infestació i proporcionar elements de gestió integrada de plagues, el seguiment del cicle de vida i la bioecologia d’aquesta plaga es van dur a terme mitjançant un recompte periòdic de poblacions a les fulles i branquetes de l’arbre del 29 de març de 2019 al 27 de març de 2020 a dues finques d’oliveres relativament llunyanes (Ain fares i Oued taria) a la wilaya de Mascara, Argèlia. Els nostres resultats van mostrar que la infestació és elevada als arbres de la granja de Oued taria. L’estudi de la bioecologia d’E. olivina va demostrar que aquesta plaga va desenvolupar dues generacions a l’any, a la primavera, en funció dels factors climàtics. Es va trobar una diferència significativa entre l’abundància relativa de les diferents etapes (ous, fase larvària 1, 2, 3, 4, i 5, femella i mascle) i les orientacions cardinals. Les orientacions del sud i del centre es van infectar més per la psyla. Les fluctuacions de les poblacions es van veure significativament afectades per les estacions a les dues finques.
Paraules clau: Bioecologia, Euphyllura olivina, Fenologia, Dinàmica poblacional, Oliverars, Plagues en oliverars
Introduction
Olive farming is a major agricultural activity in Algeria. The tree’s hardiness and plasticity allowing it to produce olives in harsh conditions (FAOSTAT, 2017). However, olive production is limited by several problems. Several insect pests, such as the olive fruit fly (Bactrocera oleae, its main pest), the olive moth (Prays oleae), the olive psyllid (Euphyllura olivina), and the black scale (Saissetia oleae) are the main causes of low productivity. These pests attack all the parts of the tree, the leaves, flowers, twigs, and fruit (El Hadrami and Nezha, 2001).
Euphyllura olivina (Costa) (Hemiptera, Sternorrhyncha, Psylloidea), native to southern Europe, is mainly found on the olive tree, Olea europaea L. (Debo et al., 2011; Triapitsyn et al., 2014), although it can also live on other Oleaceae such as Phillyrea angustifolia and P. latifolia. This species has a wide distribution, having been reported from Algeria, Austria, Corsica, France, Germany, Great Britain, Iran, Italy, Madeira, Malta, Montenegro, Morocco, Palestine, Portugal, Slovenia, Spain, Switzerland, Tunisia, and Yugoslavia. It has also been introduced into United States of America (California) (Ouvrard, 2021). It was first recorded from Algeria by Arambourg, in 1986.
Euphyllura olivina is an economic olive pest in its native regions. Its life comprises two generations (spring and autumn) or three generations (two in spring and one in autumn) (Chemseddine, 1988; Tajnari, 1992). According to Hmimina (2009), its causes most damage in spring when the developed larvae attack the flower clusters, causing their sterility (Jardak et al., 1985) and the fall of the inflorescences and fruit (Chermiti, 1983) causing losses of up to 60 % loss in some parts of the Mediterranean Basin (Zouiten and El Hadrami, 2001; Percy et al., 2012). The larvae secrete cottony flakes and honeydew which attracts ants, favouring the establishment of an ectoparasitic fungus, the sooty mold Capnodium oleaginum, which alters the photosynthesis of the tree, decreasing productivity (Arambourg and Chermiti, 1986) and thus decreasing commercial production (Chermiti, 1989; Jarraya, 2003).
Our aim was to study the population dynamics of the olive psyllid in two locations in the region of Mascara, Algeria, by means of random sampling and beating the olive tree branches (‘frappage’ method), over a 12–month period, from March 2019 to March 2020. We also aimed to: (a) determine the biological cycle of the species; (b) describe its population dynamics at various stages of development, in time (according to the seasons) and in space (two different geographic zones); and (c) determine the number of generations so as to find out the optimal period for intervention using the various control methods used against these pests in olive groves in the region.
Material and methods
Survey of the olive psyllid
The study was carried out to determine the dynamics and bioecology of the olive psyllid in olive tree groves during the growing seasons of March 2019 and March 2020. Surveys were conducted at two farms in the region of Mascara, Algeria, one in the town and the other in the other town. Ain fares is situated at 35º 28′ 47” N, 0º 14′ 39” E at an altitude of 804 m and has a surface area of 2.6 ha. The trees are over 50 years old. Oued taria is located at 35º 6’ 16” N, 0º 0’ 33” E]. The altitude is 471 m, and surface area is 3 ha. The trees are about 16 years old. Both farms have a semi–arid climate. Cultivation activity is limited to tree size and the land is ploughed in autumn. The olive trees are of the Sigoise and Chemlal variety and are planted at a distance of 8 m x 8 m on light, deep soil.
The sampling method used was that described by Vasseur and Schvester (1957), Benassy (1961) and Fabres (1979). The surveyed sites were divided into nine sampling plots including 25 olive trees. Samples were taken every 10 days. The method consists of selecting 10 trees and taking two twigs of 20 cm long and 20 leaves in each cardinal direction from the center of the tree. The samples collected were placed separately in Kraft paper bags with all information noted (date, cardinal direction, etc.).
Samples were then transferred directly to the laboratory for classifying and counting. The selected twigs and leaves were carefully examined under a binocular microscope. The different biological stages of the olive psyllids (eggs, larvae, adults) were quantified on both sides of the leaf and on the two twigs of each cardinal direction and recorded. For each stage, we quantified the total number of living individuals in order to assess the phenology or the insect over time.
Statistical analysis
The x2–test was made using the chisq.test function from R environment (R Core Team, 2020). Post hoc tests with Bonferroni correction were made using the fifer package (Fife, 2017). The significance level (ɑ) was fixed at 0.05 in this study.
Sample = (2 twigs + 20 leaves) * (4 cardinal orientation + center of the tree)/tree
Results
Population dynamics
The results of the annual fluctuation of Euphyllura olivina are shown in figure 1. The most important peaks were recorded during spring in both study areas. Olive psylla infestation started on 29/03/2019 and continued up to harvesting at 22/06/2019 with peaks of infestation on 04/05/2019 in Oued taria and on 26/04/2019 in Ain fares. The results show that the Oued taria olive grove had a higher infestation than in the Ain fares farm. Results show that the psylla infestation was affected by the region and the climate.

Fig. 1. Fluctuation of the numbers of the total population of E. olivina on the olive tree in Ain fares and Oued taria from March 2019 to March 2020. Fig. 1. Fluctuación de los valores de la población total de E. olivina en el olivo en Ain fares y Oued taria de marzo de 2019 a marzo de 2020.
The first generation in Oued taria appeared a little later than that at Ain fares. This is related to ecological conditions such as nutritional and local climatic conditions. The first generation in Ain fares began in March, coinciding with the first sap flow, and ended at the beginning of June, while the second generation started at the beginning of June and ended at the beginning of July. In Oued taria, the first generation began in late March and ended in early June, while the second generation began in early June and ended in early July. According to Kaplan and Alaserhat (2019), there is a close relationship between the ecology of E. straminae and plant phenology and climate condition.
Spatial distribution of the different life stages of E. olivina according to the orientations of the tree
According to the orientations at both study sites, we found a significant difference in the evolution of populations of the psylla E. olivina depending on the orientation of the olive branches on the trees studied (fig. 2, 3).

Fig. 2. Distribution of psyllid populations E. olivina on the olive tree according to cardinal orientations in Ain fares and in Oued taria (Mascara). Fig. 2. Distribución de las poblaciones del psílido E. olivina en el olivo según la orientación respecto a los puntos cardinales en Ain Fares y en Oued taria (Mascara).

Fig. 3. Cardinal distribution of E. olivina on the olive tree in Ain fares and Oued taria (Mascara). Fig. 3. Distribución de E. olivina respecto a los puntos cardinales en el olivo en Ain Fares y Oued Taria (Mascara).
The abundance of the different developmental stages (eggs, L1, L2, L3, L4, L5, female and male) according to the orientation was significantly different (χ2 = 49.53, df = 28, P = 0.0073) at the Ain fares farm (fig. 2). The eggs were present in high numbers on the cardinal points north, south and center, with 63 eggs/sample for each orientation. Moreover, the east and the west orientations had a lower numbers of eggs with 53 and 20 eggs, respectively. The first larval stage (L1) was about 11 larvae/sample and nine larvae/sample in the south and center, respectively. The second larval instar (L2) showed the highest numbers for the eastern orientation with an average of 19 larvae/sample and 16 larvae/sample in the south. In the third larval instar (L3) we noted a high average numbers on the north side with an average of 30 individuals per sample. For the fourth larval stage (L4), we observed that center and north directions showed the highest number of larvae with an average of 24 larvae/sample and 20 larvae/sample, respectively. The last larval instars (L5) showed respective means of 11 and 10 larvae per sample in the west and center directions. For the adult female and male, the averages were highest mainly in the south, with 35 adult females per sample and 7 male adult per sample, respectively (fig. 2). Pearson’s χ2-test showed a link between abundance of different stages of development and the cardinal orientation in the farm of Oued taria (χ2 = 50.2105, df = 28, P = 0.00613). We found the number of eggs was highest on the south side and center of the tree, with a rate of 89 eggs/sample. In the north we found about 54 eggs/sample in the north and 33 eggs/sample in the west. The first larval stage (L1) presented 13 larvae/sample in the south and 9 in the center of the tree. The second larval instar (L2) showed the highest numbers in the south with an average of 33 larvae, followed by 24 larvae/sample in the center. The third larval instar (L3) had a high number on the south side with an average of 40 larvae per sample. The fourth larval stage (L4) showed that the south of the tree had an incidence of 25 larvae/sample.The last larval instar (L5) showed that the eastern and southern directions were more infected by the larvae, with 22 and 16 larvae/sample respectively. The adult female and adult male were recorded in the south and center with numbers of 56, 34, 10, and 8, respectively. Our results showed the cumulative number of life stages of psylla according to the orientation during the season 2019 (table 1), emphasizing the positive relationships between the psylla stages, infestation and orientations. The number of L2 and L3 were higher depending on the orientation, whereas the female adult number was higher in the south, north and east. Egg numbers were very high in all the orientations. Table 1 also shows the distribution of the larval stages according to the orientation during the season 2019/2020. The number of eggs was highest in the south, center, east and north with 89, 79, 59 and 54 eggs/sample, respectively. Numbers were lowest in the west. There is a significant difference between the number of eggs and the orientation. For the second and first larval stages the numbers were highest in the south and north with 13, 33, 8 and 17 larvae/ sample respectively. The highest distribution of third, fourth and fifth instars were in the south, north and center. The difference between the larval stages and orientations was different.

Table 1. Number of olive psylla/tree recorded on infested olive in Ain fares and in Oued taria according to cardinal orientations during 2019/2020. The cumulative number in the same row with different letters significantly differs at P–value ≤ 0.05 (tests for significant differences among all pairs using x2–test; NS, not significant at P–value ≥ 0.05). Tabla 1. Número de psílidos del olivo/árbol en los olivos infestados en Ain fares y en Oued taria según la orientación cardinal durante la temporada 2019/2020. El número acumulado en la misma fila con distintas letras difiere significativamente en valores de P ≤ 0.05 (tests para diferencias significativas entre todos los pares usando el test x2; NS, no significativo para valores de P ≥ 0.05).
Monthly and seasonal distribution
The number of psylla collected varied between the two farms and season and month (fig. 4, 5). Few or no psylla was collected in winter and summer (fig. 4). In spring of 2019 to 2020 large numbers of psylla were collected in both study sites (fig. 5). Populations peaked in May. The Pearson’s chi–squared test (χ2) showed that there is a link between the presence of life stages of psylla (L1, L2, L3, L4, L5, female, male and eggs) and sampling season (χ2 = 81.0207, df = 14, P = 0). The monthly abundance and evolution of the life stage of psylla was significantly different in both study areas. The χ2–test proved the existence of relationship between the population level and months (Ain fares: χ2 = 229.37, df = 28, P < 0.001). Oued taria, a significant difference was noticed between the life stage of psylla and season (χ2 = 80.0128, df = 14, P = 0).In the same case a significant difference was observed between the life stages and the months (χ2= 196.69, df = 28, P = 0).

Fig. 4. Seasonal abundance of E. olivina in relation to life stages on olive trees in Ain fares and Oued taria (Mascara): Am, adult male; Af, adult female; E, eggs. Fig. 4. Abundancia estacional de las diferentes fases vitales de E. olivina en el olivo en Ain fares y Oued taria (Mascara): Am, machos adultos; Af, hembras adultas; E, huevos.
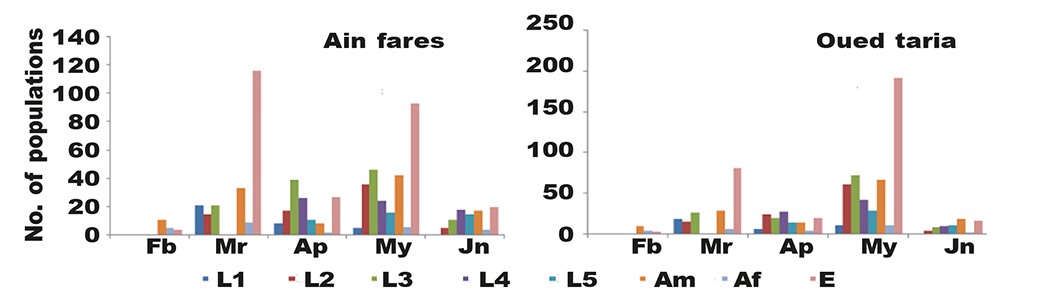
Fig. 5. Monthly evolution of the different life stages of E. olivina on olive trees in Ain fares and Oued taria (Mascara). Am, adult male; Af, adult female; E, eggs; months (Fb, February; Mr, March; Ap, April; My, May, Jn, June). Fig. 5. Evolución mensual de la abundancia de fases de desarrollo de E. olivina en el olivo en Ain fares y en Oued taria (Mascara). Am, machos adultos; Af, hembras adultas; E, huevos; meses (Fb, febrero; Mr, marzo; Ap, abril; My, mayo; Jn, junio).
In Ain fares, the results presented in table 2 and 3 showed that throughout 2019 season, the seasonal flight activity of E. olivina started at the end of February, and continued its activity throughout the season till the end of June 2019. Present study also demonstrated one peak of flight activity of E. olivina occurred at the mid of April. The cumulative number of psylla according to the season which the spring was highest season for the apparition 2019 season. Results show that the rate of psylla infestation was significantly affected by season. It was noted a higher number of eggs with 235 egg/tree in the spring followed by the L2 and L3 with 71 and 115 respectively.
A higher number of individuals were recorded during the study year from March 2019 to March 2020 in Oued taria compared (36, 105, 128, 79, 59, 117, 23, and 302) respectively for (L1, L2, L3, L4, L5, adult female and adult male). A significant difference was noted between the season and cumulative number of all the stages. The population increased gradually from the beginning of the season and decline was noted at the end of season (table 2). During this study, we have noticed the presence of eggs laid on the end of February where the numbers accelerate in March (table 3). A heavy larval infestation was noticed in the spring from March to May whereas the numbers of eggs and all larval stages were highest. Almost the population was null at the mid of June. A significant difference was registered between the larval stages number and months.

Table 2. Seasonal number of olive psylla/tree recorded on infested olive in Ain fares and in Oued taria during 2019/2020. The cumulative number in the same row with different letters significantly differs at P–value ≤ 0.05 (tests for significant differences among all pairs using x2–test; NS, not significant at P–value ≥ 0.05). Tabla 2. Número estacional de psílidos del olivo/árbol registrado en los olivos infestados en Ain fares y en Oued taria en la temporada 2019/2020. El número acumulado en la misma fila con distintas letras difiere significativamente en valores de P ≤ 0.05 (tests para diferencias significativas entre todos los pares usando el test x2; NS, no significativo en valores de P ≥ 0.05).

Table 3. Monthly number of olive psylla/tree recorded on infested olive in Ain fares and in Oued taria during 2019/2020. The cumulative number in the same row with different letters significantly differs at P–value ≤ 0.05 (tests for significant differences among all pairs using x2–test; NS, not significant at P–value ≥ 0.05). Tabla 3. Número mensual de psílidos del olivo/árbol registrado en los olivos infestados en Ain fares y en Oued taria en 2019/2010. El número acumulado en la misma fila con distintas letras difiere significativamente en valores de P ≤ 0.05 (tests para diferencias significativas entre todos los pares usando el test x2; NS, no significativo en valores de P ≥ 0.05).
Discussion
In the present study, there were no increases in populations of E. olivina in March, and the study sites had a very low population density from March to June 2019. Similarly the study of the global fluctuation of E. olivina within the study plot in Mostaganem (Algeria) by Khobzi (2019) confirmed the absence of psylla individuals on the first two sampling, and affirmed an evolution in the populations of the insect was recorded on March 23 with low abundance. Our results show the population dynamics of E. olivina from 29 March 2019 to 27 March 2020 and confirm the existence of two spring generations under the influence of climatic conditions. Most authors agree to count two generations of the insect; Mustafa (1989) in Jordan, Chemseddine (1988), Hilal et al. (1997) and Tajnari (2001) at Haouz (Morocco), Meftah et al. (2014) in Morocco, Tzanakakis (2008) in Italy and Dibo and Ksantini (2011) in Tunisia, Chafaa et al. (2017) in north–east Algeria (Batna). The population of psylla at the two farms varied, and was highest at Oued taria which was irrigated. Our results are in accordance with those of Abo–Shanab (2000) and Mesbah et al. (2019) in Egypt concerning another species of olive psyllid, E. straminae. The infestation rate was higher in the irrigated farm than in the farm with little rain, and it was seen that the activity of the insect was influenced by and linked to weather factors. The first generation had a higher population level than the second generation. These observations agree largely with those reported by Arambourg (1964), Chermiti (1989) and Ksantini (2003). Activity of the first generation of the insect appears to be influenced by the temperature and the nutritional quality of the plant, which support female fertility and larval development. Orientation with more favorable conditions seems to play a role in the development of the insect. Belhamdounia (1993) noted the preference of south orientation for larvae in the spring, and south and west orientation for adults.
The distribution of populations of E. olivina according to the season and month showed identical behavior at both study sites. Spring was the most favorable season for the development of the olive psylla, unlike E. straminea which is present in nature all year round, hibernating in the winter as adult (Kaplan and Alaserhat, 2019).
Ksantini (1986) and Chermiti (1989) observed that high temperatures in association with low relative humidity caused high mortality of eggs and larvae, and spawning even stopped in females of E. olivina. Because of the high temperatures and the slower vegetative growth of the tree, second generation adults entered summer rest from June. The insect exhibited polycyclic dynamics, showing its stability to survive from one generation to the next.
We observed an apparent difference in the life stage of psylla and season. Some other psyllid species such as Bactericera cockerelli, Cacopsylla pyricola (Förster) and Cardiaspina albitextura were reported to be intolerant to extreme weather (Blomquist and Kirkpatrick, 2002; Rakhshani and Saeedifar, 2013). Atwal et al. (1970) suggested that both high and low temperatures were detrimental to the psyllid population. However, we found a significant relationship between season and the population fluctuation of E. olivina. This phenomenon, together with different climatic conditions as shown by the rainfall from February to mid–April could explain why there was no population increase in March. Mustafa and Najjar (1985) recorded five distinct stages of ovariole formation and development. Ovarian development of the summer generation was in reproductive diapause from mid–June to mid–November. The environmental factors including temperature and photoperiod induced diapauses in adults. Adults of the spring generation showed mature eggs within a week of emergence (Mustafa and Najjar, 1985).
Conclusion and perspective
The dynamics of the psyllid E. olivina populations at the two farms in Algeria developed up to two generations per year, both in the spring and related to climatic factors. Southern and western orientations appeared to be more infested. To sum up, evolution of this psyllid is probably influenced by seasonal climatic conditions and the host plant, with spring being the most favorable season for their proliferation. Knowing the spatial distribution of an insect population helps to achieve more accurate and effective sampling and to develop a pest control management program.
Acknowledgements
We would like to thank all of those with whom we had the pleasure to work during this project.
















